
NEUROBIOLOGIA |
Cómo el sistema visual evita que el mundo desaparezca |
|
© Susana
Martínez-Conde, Stephen
L. Macknik y David
H. Hubel, 2000 smart@hms.harvard.edu Universidad de Harvard, Estados Unidos. |
RESUMEN [ABSTRACT]
Nuestro sistema visual presenta la siguiente contradicción: cuando fijamos la mirada en un objeto de interés, nuestros ojos nunca están completamente quietos. Varias veces por segundo, producimos sin darnos cuenta pequeños movimientos oculares llamados "microsacadas". Sorprendentemente, si eliminamos estos movimientos oculares en el laboratorio, nuestra percepción visual se desvanece. Si sabemos qué tipo de actividad cerebral ocurre como resultado de una microsacada, podemos saber qué tipo de actividad es importante para mantener un objeto visible. Para resolver este problema, entrenamos a macacos rhesus para que fijasen la mirada en un pequeño punto, y correlacionamos la actividad neuronal de estos monos con sus movimientos oculares. Así averiguamos que las microsacadas aumentan la actividad de las neuronas de la corteza visual primaria (área V1). Este aumento en actividad tiende a ocurrir en brotes, también llamados "ráfagas". Estas ráfagas de impulsos neuronales parecen ser el tipo de actividad más efectiva para mantener una imagen visible.
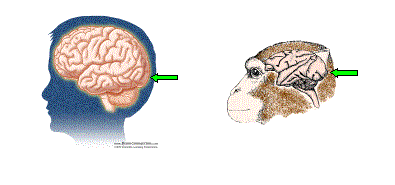
Figura 1. La información visual es procesada en la parte posterior del cerebro. Las flechas verdes indican el área V1 (o corteza visual primaria) en el humano y en el mono. (La imagen del cerebro humano ha sido cedida por BrainConnection.com; la imagen del cerebro del mono es cortesía de Patricia J. Wynne y Scientific American).
Movimientos oculares y adaptación
Nuestros sistemas sensoriales han evolucionado para detectar cambios en nuestro entorno. Una rápida reacción frente a estímulos cambiantes puede ser vital tanto para la presa como para el depredador. Cuando un estímulo no cambia a lo largo del tiempo, pierde interés para nuestros sistemas sensoriales, lo que conduce a un descenso de actividad en el cerebro. A esto se le llama adaptación neural. Un ejemplo de adaptación ocurre cuando dejamos de notar los zapatos después de tenerlos puestos durante un tiempo (a pesar de que sí notamos los zapatos en el momento de calzarnos).
El sistema visual no es una excepción. En 1952, Riggs y Ratliff y Ditchburn y Ginsborg descubrieron algo increíble: cuando se eliminan todos los movimientos oculares (por medio de aparatos especiales en el laboratorio), la percepción visual se desvanece. Cuando los ojos se mueven de nuevo, la percepción visual reaparece, de la misma manera en que Vd. es capaz de sentir los zapatos nuevamente si mueve los dedos de los pies. Los diminutos movimientos oculares que producimos involuntariamente para superar la adaptación se llaman "microsacadas".
Esta es una demostración de cómo la percepción visual desaparece durante la fijación visual.
Para hacer:
Fije su mirada en el pequeño cuadrado verde superior durante unos 30-60 segundos, o hasta que el círculo rojo desaparezca entonces mueva su mirada al cuadrado verde inferior. ¡El círculo rojo aparecerá de nuevo!Para observar:
Cuando fija su mirada con suficiente precisión, las neuronas de su sistema visual se adaptan al círculo rojo, el cual entonces desaparece. La adaptación es más efectiva si el círculo rojo resulta algo difícil de ver desde el comienzo lo que hemos tratado de hacer ajustando el brillo del círculo rojo para que sea aproximadamente igual que el brillo del fondo gris. Si el círculo le parece demasiado ocuro o demasiado claro, presione los botones "Más brillante" y/o "Más oscuro", hasta que su brillo sea igual al del fondo. Nos hemos dado cuenta de que, a veces, el círculo desaparece más fácilmente mientras su brillo está siendo ajustado activamente. Si todavía tiene dificultades para hacer funcionar esta ilusión, el modo "intermitente" puede ayudarle a igualar el brillo del círculo rojo con el fondo presione "Intermitente".Modo Intermitente:
Fije su mirada en el cuadrado verde (ahora situado arriba y en el medio). Mientras el círculo rojo parpadea contra el círculo gris, ajuste el brillo del círculo rojo (usando los botones "Más brillante" y "Más oscuro") hasta que el círculo rojo sea igual de brillante que el círculo gris. Un truco empleado por los científicos de la visión para llevar a cabo esta tarea es ajustar el brillo del círculo rojo hasta que la sensación de parpadeo del círculo se minimice. Los científicos denominan a esta técnica "fotometría del parpadeo mínimo". Cuando el círculo rojo tenga el mismo brillo que el círculo gris, presione el botón "Fijar", e intente de nuevo el test de Desvanecimiento Visual (véase "Para hacer:" más arriba) ahora el rojo será igual de brillante que el fondo gris, con lo que debería ser más fácil hacer que el círculo rojo desaparezca. El Desvanecimiento Visual es una ilusión en la que un objeto puede hacerse invisible debido a la ausencia de movimientos oculares. Este fenómeno demuestra que los pequeños movimientos de nuestros ojos son necesarios para ver con normalidad incluso cuando fijamos la mirada.FUENTE:
Martinez-Conde, S, Macknik, SL & Hubel, DH (2000) Microsaccadic Eye
Movements and Firing of Single Cells in the Striate Cortex of Macaque
Monkeys. Nature Neuroscience Vol. 3(3), pp. 251-258.
Descifrando el código neural
Puesto que las microsacadas son necesarias para mantener la visibilidad de los objetos, esto las convierte en una importante herramienta para entender el lenguaje que las neuronas usan para "hablar" entre sí: el código neural. Al correlacionar las microsacadas con la actividad neural que producen, podemos estudiar de forma indirecta el substrato neural de la visibilidad. Esto quiere decir que si sabemos qué tipos de respuestas neuronales se producen tras las microsacadas, podemos tener una idea aproximada acerca de cómo el sistema visual codifica la visibilidad de las cosas.
Las neuronas se comunican entre sí mediante impulsos eléctricos llamados "espigas" o "potenciales de acción". Si un estímulo visual aparece de repente, una neurona reaccionará produciendo más espigas que en ausencia del estímulo. Pero, ¿son todas estas espigas igualmente importantes para señalar la presencia (o ausencia) del estímulo a la neurona siguiente?
Una "ráfaga" de espigas se produce cuando una neurona dispara varias espigas consecutivamente durante un breve período de tiempo. Livingstone, Freeman y Hubel (1996) descubrieron que las ráfagas de espigas se relacionan mejor con cómo vemos la forma de los objetos que las espigas que ocurren esparcidas a lo largo del tiempo (Figura 2).

Figura 2. A y B: Cada punto representa una espiga producida por una neurona aislada del área V1 mientras el mono exploraba una escena visual que contenía un círculo blanco. La localización de cada punto muestra la posición de los ojos del mono cuando se produjo esa espiga. C y D: Los mismos datos que en A y B, ahora filtrados de forma que cada punto representa una ráfaga de espigas, y no una espiga aislada. Obsérvese que el contorno del círculo aparece solamente con ráfagas de espigas (C y D), y no con las espigas aisladas (A y B). B y D son iguales que A y C, respectivamente. El círculo se ha eliminado por claridad. [Nota 1]. (Figura reproducida con permiso de Nature Neuroscience).
Las microsacadas hacen que las neuronas disparen en ráfagas
Nos preguntamos si las microsacadas podrían también dar lugar a que las neuronas disparen en ráfagas de espigas. Para contestar a esta pregunta, llevamos a cabo un experimento similar al de la Figura 2: entrenamos monos para que fijasen la mirada en un pequeño punto, a cambio de gotas de zumo. Durante el experimento, cada mono se encuentra enfrente de un monitor de ordenador en el que se presentan los estímulos visuales. La pantalla del monitor está completamente oscura, con la excepción de un punto de fijación y de una pequeña barra de luz, colocada en el lugar más adecuado para excitar a la neurona que estemos registrando. (Decimos que la barra se encuentra en el "campo receptor" de la neurona) [Nota 2].
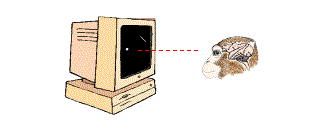
Figura 3. El mono está fijando la mirada en un pequeño punto blanco. La barra oblicua está situada en la región correspondiente al campo receptor de una neurona de V1.(La imagen del mono ha sido reproducida por cortesía de Patricia J. Wynne y Scientific American).
Mientras el mono fijaba la mirada en el punto, nosotros registramos sus movimientos oculares al tiempo que observábamos las respuestas de las neuronas individuales del área V1. La Figura 4 muestra las espigas producidas por una única neurona durante 2 segundos de registro, y también los movimientos oculares que ocurrieron en ese período. Resulta fácil ver que las microsacadas y las ráfagas de espigas están relacionadas: las microsacadas (triángulos rojos) suelen estar seguidas de una ráfaga de espigas (representada por un grupo de líneas negras en la figura), mientras que las espigas individuales tienden a ocurrir en los períodos de tiempo entre una microsacada y otra.
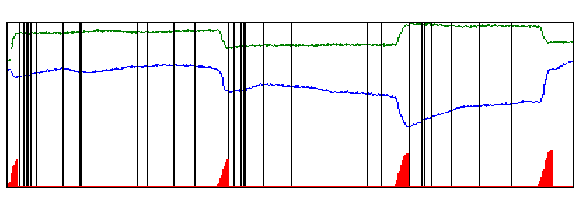 Tiempo (2 segundos)
Tiempo (2 segundos)
Figura 4. Movimientos oculares y espigas registradas durante 2 segundos. Línea verde: Posiciones oculares en el plano horizontal. Línea azul: posiciones oculares en el plano vertical. Los triángulos rojos en la parte inferior de la figura representan los movimientos oculares microsacádicos que ocurrieron durante este período. Obsérvese que aquellos cambios abruptos en el registro de los movimientos oculares (líneas verde y/o azul) se corresponden con las microsacadas (triángulos rojos). Cada línea vertical representa una espiga. Cuando dos o más espigas ocurren cercanas entre sí, decimos que este evento es una ráfaga de espigas. (Figura reproducida con permiso de Nature Neuroscience).
Cuando eliminamos la barra de luz del campo receptor de la neurona, las microsacadas no fueron seguidas de ráfagas de espigas. Esto nos indicó que las microsacadas activaban las neuronas visuales moviendo sus campos receptores dentro y fuera de la barra de luz estacionaria (lo que sería equivalente a mover la barra mientras los ojos estuvieran perfectamente inmóviles). Para que las microsacadas activen las neuronas visuales, no es suficiente que los ojos se muevan; debe haber además un estímulo visual en el campo receptor.
¿Qué es una ráfaga óptima?
En la Figura 4, puede observarse que las ráfagas de espigas que aparecen después de las 4 microsacadas son ligeramente diferentes entre sí. Algunas ráfagas contienen más espigas que otras. Incluso cuando dos ráfagas tienen el mismo número de espigas, estas espigas pueden estar espaciadas en el tiempo de forma diferente. Finalmente, incluso si dos ráfagas nos parecen exactamente iguales (mismo número de espigas espaciadas de la misma manera), pueden haber ocurrido en momentos ligeramente diferentes después de la microsacada. Puesto que las ráfagas de espigas parecen estar mejor correlacionadas con nuestra percepción visual que las espigas aisladas (Livingstone, Freeman y Hubel, 1996), quisimos averiguar qué tipos de ráfaga son más fiables en indicar que ha ocurrido una microsacada.
Para contestar a esta pregunta, observamos los diferentes tipos de ráfagas producidas por las neuronas que estudiamos, y calculamos la probabilidad de que una ráfaga determinada estuviera precedida por una microsacada. La Figura 5 muestra los resultados de este análisis para una neurona.
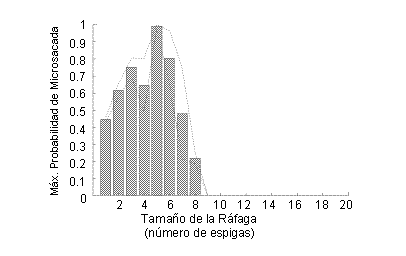
Figura 5. Probabilidad de que una microsacada preceda a ráfagas de diferentes tamaños. Una ráfaga de 1 espiga es una única espiga aislada en el tiempo. (Figura reproducida con permiso de Nature Neuroscience).
Para esta neurona, las ráfagas de 5 espigas ofrecieron la mejor correlación con microsacadas previas. Esto quiere decir que una vez que encontramos una ráfaga de 5 espigas que fuera óptima (es decir, las 5 espigas en la ráfaga se hallaban espaciadas entre sí de forma óptima y ocurrieron en el momento óptimo), pudimos estar seguros casi al 100% de que una microsacada había ocurrido antes de esa ráfaga.
Cada una de las neuronas que registramos (n=258) tuvo un tamaño de ráfaga óptimo. Para la mayoría de las neuronas, las ráfagas de 3 o más espigas fueron los mejores indicadores de microsacadas previas [Nota 3].
Conclusiones
En nuestro sistema visual, las microsacadas que producimos durante la fijación visual evitan la adaptación, al menos en parte, haciendo que las neuronas del area V1 continúen respondiendo a estímulos estacionarios. Las espigas producidas por las microsacadas no ocurren al azar, sino que están agrupadas de manera específica, habitualmente formando ráfagas de 3 o más espigas. Estas ráfagas son una clave importante para desvelar el "lenguaje" que nuestro cerebro usa para representar la visibilidad del mundo que nos rodea.
Para más información sobre este tema
Martinez-Conde S, Macknik SL and Hubel DH (2000). Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys. Nature Neuroscience 3, 251-258.
Notas
1. Las neuronas del área V1 responden de forma selectiva a una orientación específica de los bordes de los estímulos visuales (por ejemplo, una neurona determinada puede preferir las barras horizontales frente a las verticales). La neurona cuyas respuestas aparecen en la Figura 2 responde de manera máxima a una línea o borde con orientación oblicua, en este caso similar a la orientación de la línea que uniría las 10 y las 4 en el reloj. Cuando aquellas partes del círculo que corresponden a esta orientación oblicua cruzan el campo receptor visual de esta neurona, la activación es óptima.
2. Las neuronas de nuestro sistema visual responden a estímulos visuales (tales como barras de luz) que aparecen en localizaciones concretas dentro del espacio visual. La localización exacta varía de una neurona a otra. Una neurona determinada puede preferir aquellos estímulos localizados exactamente en el lugar al que estamos mirando; otra neurona puede responder solamente a aquellos estímulos localizados en una pequeña región hacia arriba y a la izquierda del lugar al que miramos. La región particular del campo visual que induce respuestas en una neurona determinada se denomina su "campo receptor". En nuestros experimentos, colocamos una pequeña barra de luz (con la orientación apropiada; ver Nota 1) en el campo receptor de la neurona que estuviéramos registrando.
3. Cuando utilizamos el término "óptimo" en este contexto nos referimos a aquellas ráfagas (y sus características) que indican una microsacada previa con una fiabilidad máxima.
Bibliografía
Ditchburn, R. W. & Ginsborg, B. L. (1952). Vision with a stabilized retinal image. Nature 170, 36-37.
Livingstone, M., S., Freeman, D. C. & Hubel, D. H (1996). Visual Responses in V1 of Freely Viewing Monkeys. Cold Spring Harb. Symp. Quant. Biol. LXI, 27-37.
Martinez-Conde S., Macknik S. L. & Hubel D. H. (2000). Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys. Nature Neuroscience 3, 251-258.
Riggs, L. A. & Ratliff, F. (1952). The effects of counteracting the normal movements of the eye. J. Opt. Soc. Am. 42, 872-873.
Punteros de Interés
Ilusiones visuales relacionadas:
Ilusión visual de desvanecimiento de un punto. http://www.exploratorium.edu/exhibits/espanol/fading_dot/index.html
Filling-in illusion. http://www.illusionworks.com/html/filling-in_illusion.html
Visual jitter illusion. http://www.visionlab.harvard.edu/Members/ikuya/html/memorandum/VisualJitter.html
Para saber más sobre el sistema visual:
Seeing, hearing and smelling the world (a report from the Howard Hughes Medical Institute). http://www.hhmi.org/senses/start.htm
BrainConnection.com. http://www.brainconnection.com/topics/?main=tophome/brain-vision
Para saber más sobre los movimientos oculares:
Mark McCourt's website. http://dinsdale.psych.ndsu.nodak.edu/homepage/Psy460/Eye%20movements/Eye%20movements.html
The painter's eye movements. http://www.physiol.ox.ac.uk/~rcm/pem
