
NEUROBIOLOGY |
How the visual system prevents the world from fading |
|
© Susana
Martínez-Conde, Stephen
L. Macknik y David
H. Hubel, 2000 smart@hms.harvard.edu Universidad de Harvard, Estados Unidos. |
[RESUMEN] ABSTRACT
Our visual system contains a built-in contradiction: when we fixate our gaze on an object of interest, our eyes are never still. Instead we produce, several times each second, small eye movements of which we are unaware, called microsaccades. Amazingly, if we eliminate these eye movements in the laboratory, our visual perception fades. By knowing the brain activity that occurs as result of a microsaccade, we can know the type of activity that is important for keeping objects visible. To address this, we trained macaque monkeys to fixate their eyes on a small spot and we correlated their neuronal activity with their eye movements. We learned that microsaccades increase the activity of neurons in the primary visual cortex (area V1). This increased activity tends to occur in clumps, also called bursts. Bursts of neuronal impulses would seem to be the type of activity most effective in sustaining a visible image.
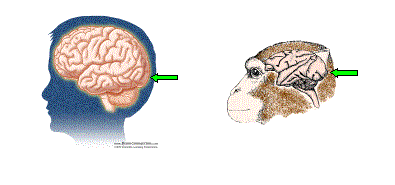
Figure 1.Visual information is processed in the back part of our brain. The green arrows indicate area V1 (the primary visual cortex) in humans and monkeys. (Human brain image courtesy of BrainConnection.com; monkey brain image courtesy of Patricia J. Wynne and Scientific American).
Eye movements and adaptation
Our sensory systems have evolved to detect changes in our environment. Reacting rapidly to changing stimuli can be vital to both prey and predators. When a stimulus does not change over time, it becomes uninteresting to our sensory systems, and this leads to decreased activity in the brain. This is called neural adaptation. An example of adaptation is when we stop noticing our shoes after having them on for a while (although we noticed them when we put them on earlier)
Our visual system is no exception. In 1952, Riggs and Ratliff and
Ditchburn and Ginsborg made the surprising discovery that when all eye
movements were eliminated (using special devices in the lab), visual
perception fades. When the eyes once again moved, visual perception
reappeared, just as you are able to notice your shoes again if you
wiggle your toes. The tiny eye movements that we make involuntarily to
overcome adaptation during visual fixation are called
microsaccades. The following is a demonstration of fading during visual fixation.
To do:
Fixate your gaze on the top green square in the upper left for approximately 30-60 seconds, or until the red spot disappears -- then move your gaze to the lower green square and the red spot will reappear!To notice:
When you fixate your gaze well enough, the neurons in your visual system will adapt to the red spot and it will disappear. To do this, it helps if the red spot is already somewhat difficult to see -- which we have tried to achieve by making it approximately the same brightness as the background, or "equiluminant". If the spot seems either too dark or too light, press the "Darker" and/or "Brighter" buttons until it is equal in brightness to the background. We have noticed that the spot sometimes disappears more readily while its brightness is actively being adjusted. If you still have difficulty getting the illusion to work, flicker-mode may help you to equate the brightnesses of the red spot with the background...press "Flicker".Flicker Mode:
Fixate your gaze on the green square (now in the middle top).As the red spot flickers against the gray spot, adjust the brightness of the red spot (using the "Darker" and "Brighter" buttons) until the red spot is the same brightness as the gray spot. A trick that visual scientists use to do this task is to set the red spot to the exact brightness that minimizes the sensation of flicker -- scientists call this technique "minimal flicker photometry". When the red spot is equal in brightness to the gray spot, press the "Fixate" button and try the Visual Adaptation test again (see "To do:" above)...now the red will be of equal brightness to the gray background and so it should be easier to make the red spot disappear.
Visual Fading is an illusion in which an object can become invisible due to lack of eye-movements. It shows that small eye-movements are necessary during normal vision -- even when we are fixating our gaze -- in order to see.
SOURCE:
Martinez-Conde, S, Macknik, SL & Hubel, DH (2000) Microsaccadic Eye
Movements and Firing of Single Cells in the Striate Cortex of Macaque
Monkeys. Nature Neuroscience Vol. 3(3), pp. 251-258.
Deciphering the neural code
Because microsaccades are necessary for keeping things visible, they become an important tool in understanding the language that neurons use to talk to each other: the neural code. By correlating microsaccades with neural activity, we can indirectly study the neural substrate of visibility. This means that if we know what kinds of neuronal responses are produced after microsaccades, we can have an idea about how the visual system encodes the visibility of things.
Neurons communicate with each other using electrical impulses called spikes or action potentials. If a visual stimulus appears suddenly, a neuron will react by producing more spikes than in the absence of the stimulus. But are all of these spikes equally important in signaling the presence (or absence) of the stimulus to the next neuron?
A burst of spikes is produced when a neuron fires several spikes in a row during a short period of time. Livingstone, Freeman and Hubel (1996) found that bursts of spikes are better related to seeing the shapes of objects than spikes that are spread out over time (Figure 2).

Figure 2. A and B: Each dot represents a spike produced by a single neuron in area V1 while a monkey is exploring a visual scene containing a white circle. The location of each dot shows the position of the monkeys eyes when that spike occurred. C and D: Same data as in A and B, but filtered so each dot now represents a burst of spikes, and not a single spike. Notice that the contour of the circle shows only for bursts of spikes (C and D) and not for single spikes (A and B). B and D are the same as A and C, but the circle has been removed for purposes of clarity. [Note 1]. (Figure reprinted with permission from Nature Neuroscience).
Microsaccades make neurons fire in bursts
We wondered if microsaccades could also cause neurons to fire in bursts of spikes. To address this question, we did an experiment similar to the one in Figure 2: we trained monkeys to fixate their gaze on a small spot in return for drops of fruit juice. During the experiment, each monkey faced a computer monitor on which visual stimuli were presented. The monitor screen was completely black, except for a fixation spot and a small bar of light, which was placed in the best location to excite the neuron we were recording from. (We say that the bar was placed in the neurons receptive field) [Note 2].
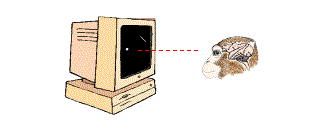
Figure 3. Monkey is fixating its gaze on a small white spot. An oblique bar has been placed in the region corresponding to the receptive field of a V1 neuron. (Monkey brain illustration courtesy of Patricia J. Wynne and Scientific American).
While each monkey stared at the spot, we recorded its eye movements and observed the responses from individual neurons in area V1. Figure 4 shows the spikes produced by a single neuron during 2 seconds of recording, and also the eye movements that occurred in that period. It is easy to see that microsaccades and bursts of spikes are related: microsaccades (red triangles) are usually followed by a burst of spikes (a group of black lines in the figure), whereas individual spikes tend to occur in the periods between microsaccades.
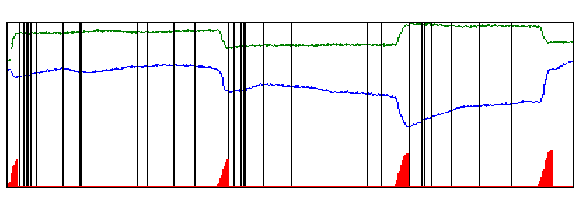
Figure 4. Eye movements and spikes collected during 2 seconds. Green trace: horizontal eye positions. Blue trace: vertical eye positions. The red triangles at the bottom of the figure represent the microsaccadic eye movements that occurred during this period. Note that abrupt changes in the eye movement record (green and/or blue traces) match microsaccades (red triangles). Each vertical black line represents a spike. When two or more spikes occur close together, we call the event a burst. (Figure reprinted with permission from Nature Neuroscience).
When we removed the light bar from the receptive field of the neuron, microsaccades were not followed by bursts. This told us that microsaccades activated visual neurons by moving the neurons receptive fields in and out of the stationary light bar (this would be equivalent to moving the bar while the eyes were held perfectly still).For microsaccades to activate visual neurons, it is not enough that the eyes move; there must also be a visual stimulus in the receptive field.
What is an optimal burst?
In Figure 4, you can see that the bursts of spikes that appear after the 4 microsaccades are slightly different from each other. Some bursts contain more spikes than others. Even when two bursts have the same number of spikes, those spikes may be spaced differently in time. Finally, even if two bursts look exactly the same (same number of spikes and same amount of spacing between them), they may happen at slightly different times following the microsaccade. Since bursts of spikes appear to be better correlated with our visual perception that single spikes are (Livingstone, Freeman and Hubel, 1996), we wanted to find out what kinds of burst were most reliable in indicating when a microsaccade happened.
To address this, we looked at the different types of bursts produced by the neurons we studied, and we calculated the probability that a given burst was preceded by a microsaccade. Figure 5 shows the results of this analysis for one neuron.
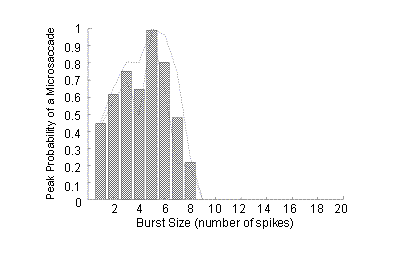
Figure 5. Probability of a microsaccade before bursts of different sizes. A burst of 1 spike is a single spike isolated in time. (Figure reprinted with permission from Nature Neuroscience).
For this neuron, bursts of 5 spikes were best correlated with preceding microsaccades. This means that once we found a burst of 5 spikes that was optimal (that is, the 5 spikes in the burst were optimally spaced apart and occurred at the optimal time), we could be almost 100% sure that a microsaccade had occurred before that burst.
Each of the neurons we recorded (n=258) had an optimum burst size. For most neurons, bursts of 3 or more spikes were the best indicators of previous microsaccades [Note 3].
Conclusions
In our visual system, the microsaccades we produce during visual fixation prevent adaptation. Microsaccades accomplish this, at least in part, by inducing V1 neurons to keep firing to stationary stimuli. Spikes provoked by microsaccades do not occur at random but are clustered in a specific manner, usually forming bursts of 3 or more spikes. These bursts are an important clue to the language our brain uses in representing the visibility of the world.
For more information on this topic
Martinez-Conde S, Macknik SL and Hubel DH (2000). Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys. Nature Neuroscience 3, 251-258.
Notes
1.Neurons in area V1 respond selectively to a specific orientation of the edges of visual stimuli (for example, a given neuron may like horizontal bars, but not vertical ones). The neuron whose responses appear in Figure 2 responds best to a line or edge in an oblique orientation, in this case roughly corresponding to a line between 10 oclock and 4 oclock. This neuron is thus most active when its visual receptive field is crossed by parts of the circle that match this oblique orientation.
2.Neurons in our visual system respond to visual stimuli (such as bars of light) that appear in particular locations in the visual space, the exact location varying from neuron to neuron. A given neuron may like stimuli located just in the specific place we are looking at; another neuron may only like stimuli located in a small region up and to the left of where we are looking. The particular location in the visual field that induces responses in a given neuron is called its receptive field. In our experiments, we placed a small bar of light (with the appropriate orientation; see Note 1) in the receptive field of the neuron we were recording from.
3.When we use the term "optimum" in this context, we refer to those bursts (and their characteristics) that indicate a previous microsaccade with maximum reliability.
References
Ditchburn, R. W. & Ginsborg, B. L. (1952). Vision with a stabilized retinal image. Nature 170, 36-37.
Livingstone, M., S., Freeman, D. C. & Hubel, D. H (1996). Visual Responses in V1 of Freely Viewing Monkeys. Cold Spring Harb. Symp. Quant. Biol. LXI, 27-37.
Martinez-Conde S., Macknik S. L. & Hubel D. H. (2000). Microsaccadic eye movements and firing of single cells in the striate cortex of macaque monkeys. Nature Neuroscience 3, 251-258.
Riggs, L. A. & Ratliff, F. (1952). The effects of counteracting the normal movements of the eye. J. Opt. Soc. Am. 42, 872-873.
Interesting Links
Related Visual Illusions:
Fading-dot illusion. http://www.exploratorium.edu/exhibits/espanol/fading_dot/index.html
Filling-in illusion. http://www.illusionworks.com/html/filling-in_illusion.html
Visual jitter illusion. http://www.visionlab.harvard.edu/Members/ikuya/html/memorandum/VisualJitter.html
To learn more about the visual system:
Seeing, hearing and smelling the world (a report from the Howard Hughes Medical Institute). http://www.hhmi.org/senses/start.htm
BrainConnection.com. http://www.brainconnection.com/topics/?main=tophome/brain-vision
To learn more about eye movements:
Mark McCourt's website. http://dinsdale.psych.ndsu.nodak.edu/homepage/Psy460/Eye%20movements/Eye%20movements.html
The painter's eye movements. http://www.physiol.ox.ac.uk/~rcm/pem
Glossary
Adaptation: A decrease in neural activity in response to steady stimulation over time.
Burst of neuronal impulses: Sudden increment in the activity of a nerve cell during a brief period of time.
Cortex: Most external part of our brain. It processes complex information and is responsible for our cognitive abilities.
Neuron: Excitable nerve cell specialized in receiving, processing, and sending electrochemical signals.
Visual receptive field: Region in the visual world that is seen by a given neuron.
Visual system: The part of our nervous system specialized in receiving and processing visual information.
Biographies
Susana Martínez-Conde was born in A Coruña, Spain. She received her Ph.D. in 1996 from the University of Santiago de Compostela, where she worked with Javier Cudeiro and Carlos Acuña. In 1997 she joined David Hubels laboratory at Harvard Medical School, where she has been since. For further information, visit her web page at http://cortex.med.harvard.edu/~martinez-conde
Stephen Macknik is from the island of Maui, Hawaii. He received his Ph.D. in Neurobiology from Harvard University in 1996, where he worked with Margaret Livingstone. He has since been in the laboratory of David Hubel, at Harvard Medical School. For more information, contact his web page at: http://cortex.med.harvard.edu/~macknik
David H. Hubel was born in Ontario, Canada. He graduated from McGill University in 1947, with a BSc in Mathematics and Physics, and received his M.D. from McGill Medical School in 1951. In 1958, he began collaborating with Torsten Wiesel, first at John Hopkins and then at Harvard Medical school, where he has been ever since. In 1981, he shared the Nobel Prize in Physiology or Medicine with Torsten Wiesel and Roger Sperry.
