
NEUROBIOLOGIA |
Cómo darle forma a nuestro cerebro: Moldeando la corteza cerebral |
|
©
Guillermina López-Bendito
,
2004
guillermina.lopez-bendito@anat.ox.ac.uk |
RESUMEN[ABSTRACT]
El cerebro de mamíferos es el órgano más fascinante y complejo estudiado jamás. Alrededor de 100 billones de células, llamadas neuronas, se organizan formando una inmensa red de conexiones mediante la cual la información procedente del exterior se procesa. Interesantemente, muchas de estas conexiones se realizan en etapas muy tempranas, embrionarias, de nuestro desarrollo. El balance entre información intrínseca (genética) y extrínsica (proveniente de estímulos del exterior) parece jugar un papel crucial en el correcto desarrollo de nuestro cerebro. Una de las más laboriosas e importantes conexiones que realiza nuestro cerebro es la que ocurre entre el núcleo de relevo de toda la información sensorial, el tálamo, y la región donde la mayor parte de esa información es procesada, la corteza cerebral. Localizadas a varias leguas de distancia, estas dos regiones tienen que establecer una conexión permanente de manera controlada y precisa. Al mismo tiempo, la corteza cerebral se divide en distintas áreas anatómicas y funcionales encargadas de procesar distintas modalidades sensoriales y motoras. En este proceso de regionalización parecen participar también factores genéticos y epigenéticos, estos últimos provenientes en su mayoría de la información desde el tálamo. Algunos de los mecanismos por los que ocurren estos magníficos eventos, la regionalización de la corteza cerebral y el desarrollo de las conexiones tálamo-corticales, son discutidos a continuación.
La complejidad anatómica del tejido nervioso no fue apreciada hasta la invención del microscopio compuesto. Hasta mediados del siglo XVIII, el tejido nervioso se consideraba un tejido glandular implicado en el transporte de fluido secretado por el cerebro hacia el resto del cuerpo mediante diferentes conductos o nervios. Fue a finales del siglo XIX cuando la histología del sistema nervioso central (SNC) empezó a describirse de forma más precisa a través de los estudios de Camillo Golgi y Santiago Ramón y Cajal. Fue éste ultimo quien demostró, en contra de las teorías de aquella época, que el sistema nervioso está constituído por elementos individuales denominados neuronas. Actualmente, sabemos que el sistema nervioso está constituido por dos grandes tipos de células: las neuronas y las células gliales. Las neuronas cumplen la función de recibir e integrar información y de enviar señales a otros tipos de células excitables a través de contactos sinápticos.
Organización del sistema nervioso
El sistema nervioso central (SNC), el cual es bilateral y esencialmente simétrico, consta de seis grandes regiones: la médula espinal, la médula oblongata, el puente y cerebelo, el mesencéfalo, el diencéfalo, y los hemisferios cerebrales. Los hemisferios cerebrales representan lejos la región más grande del cerebro. Están formados por la corteza cerebral, la sustancia blanca, y tres núcleos profundos: los ganglios basales, la formación hipocampal, y la amígdala. En el curso del último siglo y medio, mediante el empleo de nuevas técnicas de análisis e imagen, el manto cortical se ha podido dividir en alrededor de 50 a 100 áreas con distintas características funcionales y estructurales (Figura 1).
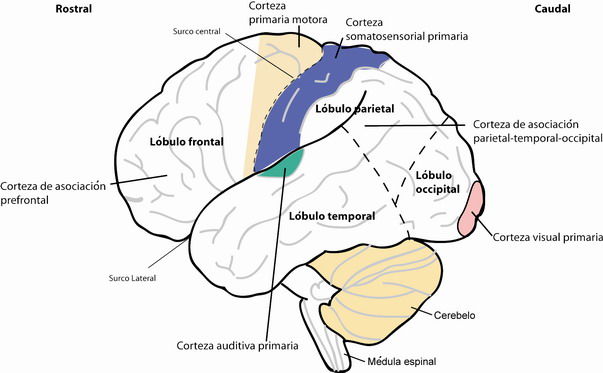
Figura 1. Regionalización de la corteza cerebral humana ilustrando sus áreas más relevantes, las cortezas primarias somatosensorial, motora, auditiva y visual.
Quizás el más famoso de los mapas que describen la regionalización cortical fue generado por Korbinian Brodmann en la primera década del siglo XX, aunque interpretaciones alternativas han sido posteriormente publicadas. Los mapas de regionalización cortical representan una guía indispensable y fundamental a la hora de comprender la citoarquitectura (ver glosario) y funcionamento del SNC. Como ejemplo de ello tenemos el núcleo talámico o simplemente tálamo. El tálamo es el núcleo de relevo de la mayor parte de la información sensorial que nos llega desde el exterior y que acaba en la corteza cerebral para ser procesada. Virtualmente todo el tálamo projecta topográficamente (ver glosario) al área sensorial correspondiente del manto cortical, el cual manda conexiones de vuelta al correspondiente núcleo talámico (Figura 2 y Figura 4). De una manera u otra, la información de cada modalidad sensorial alcanza su respectiva área en la corteza cortical, y ésta manda de vuelta conexiones e información al sistema motor.
Cada hemisferio de la corteza cerebral se caracteriza por procesar información procedente del lado contralateral del cuerpo. La información sensorial o motora procedente del lado izquierdo o derecho del cuerpo entra en la médula espinal y cruza al lado opuesto del sistema nervioso antes de pasar por la corteza cerebral donde es procesada.
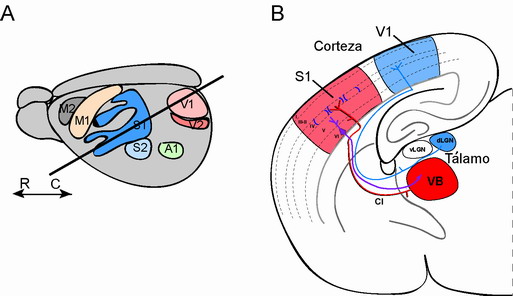
Figura 2. Diagrama que ilustra las más relevantes áreas de la corteza cerebral en roedores (A). Si se realiza un corte rostro (R)-caudal (C) con un ángulo de ~ 60 grados se expone la via de conexiones desde los núcleos talámicos hasta la corteza cerebral (B). Cada nucleo talámico recibe una determinada información sensorial del exterior. Por ejemplo, información sensorial llega al complejo ventrobasal (VB, rojo) del tálamo o información visual al núcleo geniculado lateral (LGN, azul). A partir de aquí la información se envía a la corteza correspondiente, sea ésta somatosensorial (S1) o visual (V1), respectivamente. CI, cápsula interna.
Formación del patrón cortical
La corteza cerebral, región del cerebro única en mamíferos, comprende distintas subregiones definidas anatómicamente por el número de capas (por ejemplo, la neocorteza contiene seis capas (I a VI) mientras que el hipocampo contiene sólo tres) y, como mencionamos anteriormente, por un gran número de áreas funcionales definidasmás de cincuenta en la neocorteza humana. Una de las dificultades en entender cómo se desarrollan las distintas áreas corticales surge de que en pequeños mamíferos como el ratón y la rata, la neocorteza es virtualmente uniforme en estructura y citoarquitectura. Además, como mencionaremos en detalle más adelante, son sólo unos pocos los genes conocidos que se expresan en etapas tempranas del desarrollo y que puedan ser elegidos como candidatos en el control del patrón de áreas corticales. Por lo tanto, por mucho tiempo se pensó que este patrón cortical era impuesto en la neocorteza únicamente mediante factores extrínsicos, principalmente mediante las aferencias talámicas que alcanzan la corteza manteniendo las proyecciones específicas a cada área cortical. Sin embargo, recientes evidencias apuntan a que el neuroepitelio cortical (área de la corteza donde se originan la mayor parte de las neuronas) adquiere el patrón regional antes de que la inervación procedente del tálamo alcance su diana en la corteza. Recientes estudios apuntan a que genes específicos y/o factores de transcripción (ver glosario) se expresan de manera gradual a lo largo del neuroepitelio cortical, siendo su expresión a su vez controlada por concentraciones locales de factores difusibles. Por ejemplo, los genes homeóticos Emx2 y Pax6 se expresan en gradiente opuesto uno del otro pero a lo largo del eje anterioposterior de la corteza (Figura 3), mientras que factores de transcripción como Lef1 se expresan gradualmente en el eje medio-lateral. Mediante el uso de ratones transgénicos (ver glosario), recientemente se ha puesto de manifiesto que los factores de transcripción Emx2 y Pax6 están directamente relacionados con la formación de las áreas corticales (O'Leary & Nakagawa, 2002). Por tanto, estos factores de transcripción son responsables directamente del gradiente de expresión normal en la corteza, mientras que la deficiencia de estos genes causa disminución o amplificación de áreas (Figura 3).
¿Cómo los axones talámicos afectan la formación de áreas en la corteza?
Como ya mencionamos anteriormente, los principales núcleos del tálamo dorsal envían proyecciones a la neocorteza siguiendo una topografía específica. Numerosos estudios han puesto de manifiesto que este input talámico es definitorio para el correcto desarrollo no sólo anatómico sino también funcional de las distintas áreas corticales (O'Leary, 1989; Rakic, 1988). Así, la especificidad de la proyección talámica por cada área cortical se mantiene a lo largo de su desarrollo, estando la posterior diferenciación de las
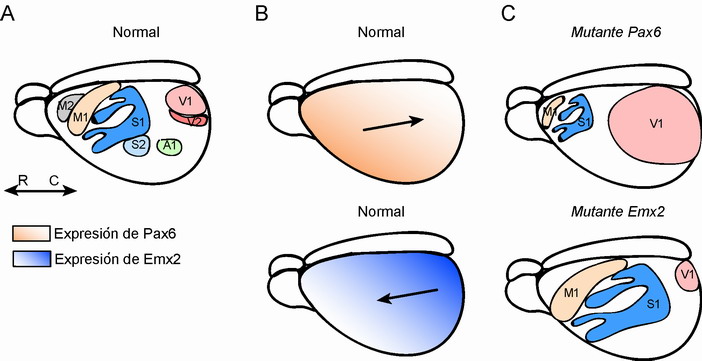
Figura 3. Modelo de patrón cortical en la corteza cerebral en roedores (A). Factores de transcripción como Emx2 y Pax6 se expresan en gradiente opuesto y en etapas muy tempranas del desarrollo embrionario (B). En colores naranja y azul se muestra el gradiente de expresión de Pax6 y Emx2, respectivamente. Las flechas indican el eje rostro-caudal (B, arriba) o caudo-rostral (B, abajo). La carencia de estos genes ha demostrado tener un fuerte impacto en el desarrollo de las diferentes áreas corticales (C). Por ejemplo, la ausencia del gen Pax6 produce una expansión de áreas caudales del la corteza como el área visual primaria (V1). En el caso del gen Emx2 su carencia produce la expansión de áreas rostrales como las áreas motora primaria (M1) o somatosensorial primaria (S1).
capas corticales directamente relacionada con el refinamiento de los axones talámicos a nivel cortical. Aunque algunos genes se expresen tempranamente en la corteza cerebral y sean capaces por sí solos de establecer un cierto nivel de regionalización, el input talámico podría ser requerido para el mantenimiento de estas diferencias. Actualmente, existe un gran debate acerca de si el input talámico es suficiente o no para crear la regionalizacion en la corteza, o si por el contrario la información intrínseca (genética) es suficiente en mediar este hecho. Huffman y colaboradores realizaron en 1999 un experimento clave con intención de clarificar este punto (Huffman et al, 1999). El experimento consistió en la ablación de una gran parte de la corteza cerebral antes de que la invasión talámica tuviese lugar, delimitando considerablemente la diana para los axones talámicos. Interesantemente, la corteza cerebral restante desarrolló las áreas principales, somatosensorial auditiva y visual, siguiendo un patrón espacial similar al que ocurre en animales normales. Además, la relación topográfia por parte de los núcleos talámicos y de sus proyecciones al área de corteza correspondiente estaba igualmente conservada.
Los resultados expuestos anteriormente apoyan el hecho de que los axones talámicos se encuentran en una posición ideal para proveer a la corteza de información necesaria para su regionalización. Su acceso al mundo exterior por medio de los órganos sensoriales, la especificidad de sus proyecciones, y su contacto temprano con las células diana en la capa IV son componentes importantes de esta hipótesis.
Desarrollo de la proyeccion talámica: Factores implicados
El desarrollo de la corteza cerebral y del tálamo ocurre sincrónicamente. La mayoría de las células o neuronas del tálamo se generan en la rata entre los días embrionarios 13 y 19 (siendo el periodo gestacional de 22 días), el cual coincide con el periodo de desarrollo neuronal en la corteza. Los axones procedentes del tálamo inician su largo recorrido en este período teniendo que cruzar y navegar a través de distintas regiones del cerebro hasta alcanzar su destino (Figura 4). Pero, ¿cómo saben los axones talámicos dónde tienen que ir y dónde tienen que detenerse?. En los últimos años, muchos están siendo los esfuerzos encaminados a comprender los mecanismos implicados en este proceso. Al igual que ocurre en la corteza, genes y factores de transcripción se expresan también durante su desarrollo en regiones navegadas por los axones talámicos. Experimentos recientes sugieren que la guía de axones talámicos ocurre en parte mediante la expresión y función de estos genes y/o factores de transcripción (López-Bendito & Molnár, 2003). Estudios en detalle en ratones mutantes o deficientes en algunos de estos genes o factores de transcripción han puesto de manifiesto este hecho. La falta de expresión y, por tanto, de función de alguno de estos genes causa anomalías en el desarrollo, recorrido y término de la proyección talámica a la corteza. La carencia de, por ejemplo, los genes Pax6 o Gbx2, expresados en el neuroepitelio cortical y en el tálamo dorsal, respectivamente, tiene como consecuencia la total ausencia de axones talámicos en la corteza.
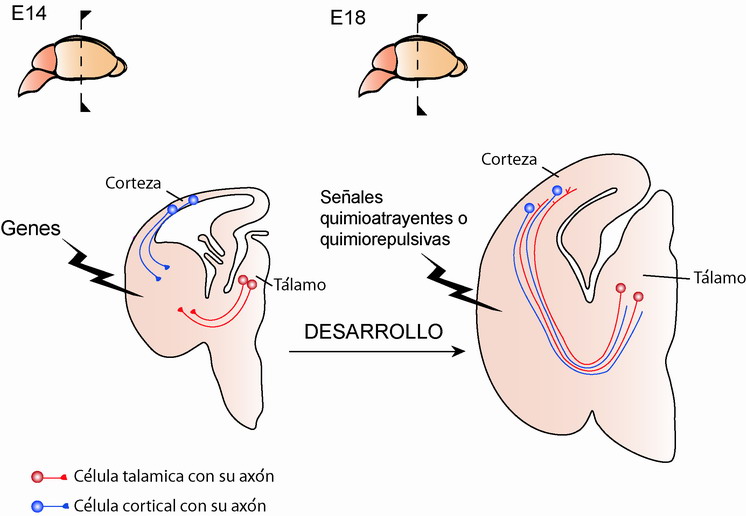
Figura 4. Desarrollo del sistema tálamo-cortical en roedores. Las neuronas del tálamo y corteza envían sus axones o proyecciones en estadíos muy tempranos del desarrollo embrionario (E14). Durante su largo viaje muchos son los factores conocidos involucrados en controlar la correcta conexión entre estas dos grandes regiones del cerebro.
Otros factores que parece estar implicados en la guía de la proyección talámica hasta su diana en la capa IV, es la liberación desde la corteza de factores con función repulsora o atrayente (Figura 4). Ejemplo de ello son los factores llamados Efrinas y Neurotrofinas (NT), moléculas y factores de crecimiento que se expresan y liberan por células de la corteza y que actúan sobre los axones talámicos permitiendo y refinando su entrada en el área cortical adecuada. En ratones mutantes en los que la expresión de la neurotrofina 3 (NT-3) en la corteza ha sido interrumpida, se produce una pérdida masiva de proyección talámica en regiones concretas de la corteza, como en la corteza visual. Adicionalmente, receptores para estas neurotrofinas, como el receptor p75, se expresan en forma de gradiente en la corteza. En el ratón mutante en el cual la expresión de p75 se encuentra inhibida, el input talamocortical en la corteza visual está considerablemente reducido. Los resultados expuestos anteriormente apuntan a que las neurotrofinas y sus receptores juegan un papel importante para que los axones talámicos alcancen su correcta diana en la corteza.
Llegada a la corteza
En mamíferos, las fibras o axones talámicos ingresan a la corteza antes de que las células diana de capa IV se hayan formado, teniendo que esperar dos o tres días a que este evento ocurra. Otro hecho importante digno de mencionar es que el desarrollo de la conectividad entre el tálamo y la corteza ocurre antes de que las sensaciones del exterior alcancen el tálamo dorsal. Esto es, el cerebro se prepara en sus conexiones para la llegada de la información desde el exterior. Se ha sugerido en los últimos años que mientras los axones talámicos esperan a las puertas de la corteza para entrar, invadir y establecer contacto con sus células diana, ellos establecen pequeñas conversaciones con los centinelas. Llamamos centinelas a un grupo de células que se localizan en la parte más profunda de la corteza, denominadas células de subplaca, capa por la que los axones talámicos tienen que cruzar (Figura 5). Estas conversaciones consisten en señales eléctricas, mediadas por estrechos contactos entre los axones y las células de subplaca, que parece que son importantes en darle la señal de entrada a los axones talámicos en la corteza.
Una vez que las células de capa IV se localizan en su posición final en la corteza, los axones talámicos cruzan la subplaca y se dirijen hacia ésta siguiendo una dirección perpendicular a la superficie de la corteza; esta invasión ocurre hacia el primer día postnatal (Figura 5). Pero, ¿cómo saben los axones cuándo tienen que parar dado que han llegado al final de su recorrido?. Recientes estudios han sugerido la existencia de una señal de parada localizada en la corteza, y que está implicada en determinar la especificidad de los axones talámicos en detenerse en la capa apropiada. Estudios in vitro indican que los axones talámicos son capaces de reconocer sus células diana de entre todas las demás. En experimentos de co-cultivo donde explantes de corteza y tálamo se confrontan directamente, las conexiones tálamo-corticales se forman siguiendo la misma especificidad de capa que realizan in vivo. La existencia de una señal de parada se ha demostrado más directamente usando técnicas de captación de imágenes en tiempo real en cultivos celulares. Mediante esta técnica se puede seguir la trayectoria de axones in vitro (Yamamoto et al, 1997). En estos experimentos se realizaron explantes en los que el tálamo se coloca en diferentes posiciones relativas a la corteza. En estas condiciones, los axones talámicos siempre paran y realizan sus conexiones en capa IV, independientemente de su ángulo de entrada. Estos resultados sugieren que la señal o señales de parada se encuantran en la corteza posiblemente siguiendo un gradiente de expresión, siendo éste más intenso en la capa diana, la capa IV. La identidad de esta señal todavía no se ha descubierto pero existen algunos candidatos. Moléculas de la matriz extracelular o localizadas en la superficie de determinadas neuronas, incluyendo algunas Efrinas, semaforinas o caderinas, que se expresan siguiendo un patrón específico de capa, podrían tener una función determinante en este hecho.
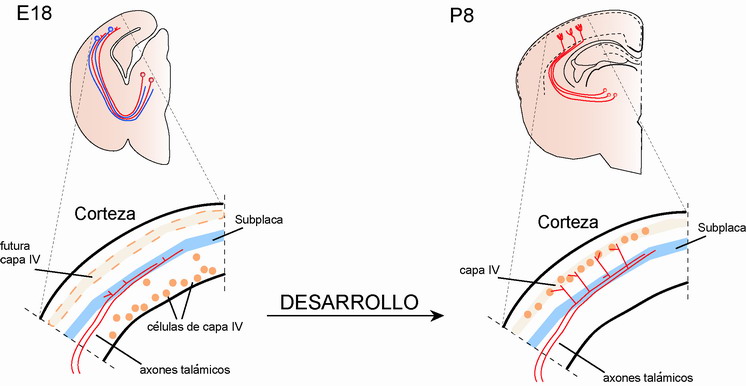
Figura 5. Innervación por los axones talámicos de la capa IV cortical. Cuando los axones talámicos llegan a la corteza, sus células diana no han llegado a su destino, por lo que tienen que esperar en la subplaca (E18). Se ha sugerido que los axones talámicos establecen señales eléctricas con las células de subplaca durante este período. Este contacto parece ser importante para que los axones invadan la corteza en el momento y región adecuada. Una vez que las células de capa IV alcanzan su posición final, los axones talámicos la invaden estableciendo su conexión final (P8).
Conclusiones
Como hemos descrito en este artículo, el desarrollo cortical implica el moldeamiento del epitelio cortical en discretas áreas con aferencias, eferencias y redes de conexiones específicas. La interacción e intercambio de información entre la corteza y el tálamo desde muy temprano en el desarrollo parece jugar un papel importante en este hecho. Durante los últimos años se ha realizado un progreso importante hacia la comprensión de cuáles son los mecanismos implicados en el desarrollo del sistema tálamo-cortical y de su interacción temprana con el circuito cortical. Estos avances se han realizado mayoritariamente estudiando ratones mutantes deficientes en genes o factores de transcripción que se expresan en distintas regiones del cerebro. Ahora, nuestra tarea es la de interpretar estas observaciones y establecer la relación causa-efecto entre estos genes y factores expresados, y el patrón talámico y cortical establecido.
Bibliografía
O'Leary DD, Nakagawa Y. (2002) Patterning centers, regulatory genes and extrinsic mechanisms controlling arealization of the neocortex. Curr Opin Neurobiol. 12:14-25. Review
O'Leary DD (1989) Do cortical areas emerge from a protocortex?. Trends Neurosci 12:400406.
Rakic P (1988) Specification of cerebral cortical areas. Science 241: 170176
Huffman K.J, Molnár Z, Van Dellen A, Kahn D.M, Blakemore C, Krubitzer L (1999) Formation of cortical fields on a reduced cortical sheet. J. Neurosci. 19:99399952
López-Bendito G, Molnár Z (2003) Thalamocortical development: how are we going to get there? Nat Rev Neurosci. 4:276-89. Review.
Yamamoto N, Higashi S, Toyama K (1997) Stop and branch behaviors of geniculocortical axons: a time-lapse study in organotypic cocultures. J Neurosci. 17:3653-63.
Glosario
1. Factores de transcripción: Término que se aplica a cualquier proteína requerida en iniciar o regular la transcripcion del ADN en eucariotas. Esto incluye, genes que regulan proteínas y factores de transcripción en general.
2. Ratones transgénicos: Ratones que han sido modificados genéticamente mediante la inserción o delección de un o varios genes específicos. Los genes que son insertados o modificados se denominan transgenes. En casos especiales en los que un o varios genes de interés son inactivados al animal resultante se denomina ratón knock-out.
3. Citoarquitectura: estructura de una región del sistema nervioso caracterizada por las relaciones entre células y sus ramificaciones y conexiones.
4. Projección topográfica: proyección de axones de una región del sistema nerviosa a otra diferente y distante con preservación de relaciones laterales de neuronas adyacentes.
5. Cultivos celulares: células de un órgano o tejido son disociadas usando diversas técnicas las unas de las otras y mantenidas en condiciones artificiales (o de cultivo). Así mantenidas, neuronas pueden adherirse al nuevo sustrato (vidrio o plástico), crecer y establecer nuevas conexiones.