
BOTANICA |
Comunicación intercelular a distancia a través del floema en plantas |
|
©
María de Jesús Elisa León Ramírez,
Beatriz Xoconostle Cázares,
Roberto Ruiz Medrano
,
2004
rmedrano@enigma.red.cinvestav.mx |
RESUMEN[ABSTRACT]
En muchos casos, la respuesta a diferentes estímulos ambientales en plantas ocurre en un tejido diferente al que percibe el estímulo, como en la respuesta al ataque de patógenos y la floración. En el primer caso, se establece una respuesta de defensa en tejidos no infectados, mientras que en el segundo el ápice del brote se transforma en estructura reproductiva como respuesta a señales producidas en las hojas que han percibido el estímulo, y que son transportadas a otros tejidos a través del floema. Este es el conjunto de conductos vasculares que distribuyen nutrientes producto de la fotosíntesis a toda la planta. Se han propuesto hormonas vegetales clásicas y otras moléculas relativamente pequeñas como señales, pero evidencias más recientes sugieren que proteínas y ácidos nucleicos podrían también serlo, considerando el transporte de virus dentro de plantas por floema y la propagación del silenciamiento génico postranscripcional.
El transporte de nutrientes en plantas ocurre a través del tejido vascular, extensa red de vasos que derivan de células altamente modificadas. Está constituido por el xilema, que transporta tanto agua como nutrientes minerales de la raíz al resto de la planta, y el floema, que distribuye azúcares, aminoácidos y otros nutrientes de los tejidos fotosintéticamente activos a los que no son capaces de fotosintetizar (Figura 1) (Taiz & Zeiger, 2002). En su madurez algunas células del floema pierden la mayor parte de sus orgánulos (incluido el núcleo), pero mantienen la integridad de su membrana celular, mientras que en el xilema sufren un proceso análogo al de muerte celular programada, resultando en vasos con una pared celular reforzada. En ambos tejidos ocurre el libre movimiento de estructuras tan grandes como orgánulos o patógenos microbianos. La función de distribución de nutrientes del tejido vascular en plantas es esencial, pero algunas evidencias sugieren que el papel del tejido vascular en el desarrollo de plantas es más activo, particularmente en el caso del floema.
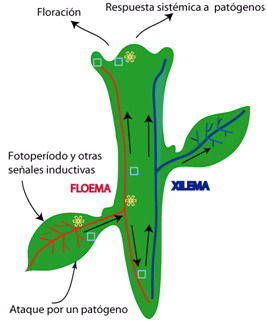
Figura 1. Esquema que representa al floema (rojo) y xilema (azul). Las flechas indican la dirección del flujo de ambos conductos; así, mientras que la savia del xilema fluye de las raíces al resto de la planta, la savia del floema fluye de las hojas fotosintéticas al resto de la planta. Se recalca el papel del floema como vía de transporte de señales que inducen fenómenos tales como la floración y establecimiento de una respuesta de defensa contra patógenos en el meristemo apical. El asterisco indica la señal, transportada desde hojas maduras e inducida tanto por un programa genético como por fotoperíodo y otras señales, que induce la floración. El cuadrado pequeño corresponde a la señal que induce la respuesta defensa contra patógenos en tejidos no expuestos a éstos. Esta señal también es transportada del sitio original de la infección al meristemo apical, así como a otros tejidos.
El floema
El floema consta en su madurez de dos tipos celulares, la célula acompañante y el elemento criboso, interconectadas por plasmodesmos (Figura 2). Mientras que la célula acompañante mantiene su integridad y elevada actividad metabólica, el elemento criboso en su madurez pierde el núcleo y orgánulos en general, y las separaciones con los vecinos anticlinales se transforman en placas cribosas que permiten el libre tránsito de moléculas de gran tamaño. Sin embargo, en contraste con el xilema su membrana plasmática permanece funcional. Además, mientras que el xilema es continuo con el espacio extracelular de toda la planta, el floema es continuo con el citoplasma de casi todas sus células. Los azúcares son exportados del mesófilo, en donde se fija carbono, hasta el floema, ya sea a través de los plasmodesmos o por el espacio extracelular. Una vez en el floema, los nutrientes son transportados rápidamente a los tejidos consumidores, en donde requieren ser "descargados" a las células circundantes. El floema también distribuye señales que intercomunican tejidos distantes, un papel que es evidente en la inducción de la floración, la respuesta de defensa contra patógenos y otros más que son descritos a continuación.
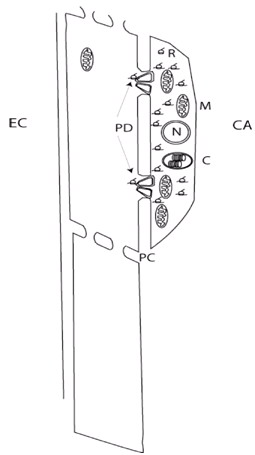
Figura 2. Diagrama esquemático del complejo célula acompañante (CA) y elemento criboso (EC) del floema. PD denota los plasmodesmos modificados que comunican estos dos tipos celulares. En esta ilustración el número relativo de orgánulos tales como mitocondrias, ribosomas y cloroplastos indica su mayor concentración en la célula acompañante con respecto al elemento criboso (o su ausencia, como en el caso del núcleo). N, núcleo; M, mitocondria; C, cloroplasto; R, ribosoma; PC, placa cribosa.
Inducción de la floración
En este fenómeno, el meristemo apical del brote da lugar a otro grupo de células precursoras de la flor como respuesta a una señal que proviene de las hojas (Figura 1). Dicha señal ha sido denominada florígeno, si bien este término no es muy aceptado. Las hojas reciben el estímulo para florecer (una combinación de condiciones ambientales y un programa genético) (Colasanti & Sundaresan, 2000; Blázquez et al., 2003), y en respuesta producen una señal que es transportada por el floema al meristemo apical, en donde se origina el primordio floral. Este fenómeno puede ocurrir en ausencia de estímulos ambientales. Se ignora la naturaleza de la señal, o señales, pero se han sugerido azúcares, hormonas vegetales, péptidos y otras moléculas difusibles. En maíz, plantas cuyo gen indeterminate 1 (id1) es defectuoso ignoran el estímulo que en circunstancias ordinarias conduciría a la floración. Este gen es activo en células que rodean al floema de hojas maduras e inmaduras. Más aun, codifica un regulador transcripcional, por lo que es tentador especular que está involucrado en la producción de la elusiva señal florigénica (Colasanti & Sundaresan, 2000).
Resistencia sistémica adquirida al ataque de patógenos
Como respuesta al ataque de patógenos, se activan numerosos genes cuyos productos degradan la pared celular de bacterias u hongos, destruyen células infectadas, etc. Esta inducción no ocurre sólo en el tejido inicialmente infectado, sino en hojas y otros tejidos expuestos al patógeno gracias a señales que son transportadas a través del floema (Ruiz-Medrano et al., 2001). El ácido salicílico es capaz de inducir genes de defensa en hojas no infectadas, si bien algunos trabajos sugieren que éste es necesario para establecer la respuesta de defensa en hojas locales y sistémicas, pero no es la señal móvil misma (Ryals et al., 1995). Experimentos recientes han demostrado en Arabidopsis thaliana que una proteína de unión a lípidos es necesaria para el establecimiento de la resistencia sistémica adquirida, lo que podría sugerir que las señales móviles son de naturaleza lipídica (Maldonado et al., 2003). Por otra parte, daño mecánico por insectos activa genes de inhibidores de proteasas, altamente tóxicos para éstos. Señales transportadas en el floema inducen estos genes también en hojas intactas. En algunas especies, se ha encontrado que un péptido sintetizado en la célula acompañante, la sistemina, es la señal transportada (Ryan y Pearce, 1998).
Respuesta a déficit hídrico
Como respuesta déficit hídrico en el suelo, las plantas producen una molécula reguladora del crecimiento, el ácido abscísico (ABA). Este es transportado por el xilema desde la raíz a hojas maduras en donde induce el cierre de los estomas, evitando una mayor pérdida de agua (Ruiz-Medrano et al., 2001). Otra respuesta observada es un menor crecimiento de la parte aérea de la planta con relación a la raíz; asimismo, el meristemo vegetativo se transforma en uno floral más rápidamente. Así la planta acelera eventos tales como la floración, a la vez que concentra sus recursos en la producción de raíces. Reguladores específicos del ciclo celular se inducen como respuesta a ABA, por lo que es posible que esta fitohormona induzca la síntesis de una o más señales secundarias en las hojas que a su vez son transportadas a través del floema al resto de la planta, particularmente al meristemo apical del brote y al de la raíz. De esta forma, el primero permanece latente mientras que las células de los meristemos de la raíz continúan desarrollándose.
Silenciamiento génico postranscripcional
El silenciamiento génico postrancripcional consiste en la degradación específica de ARN de virus en tejidos no infectados, o bien, de transgenes particularmente activos y que son muy semejantes en secuencia a genes endógenos (Vance & Vaucheret, 2001). Se considera que este fenómeno es en realidad un mecanismo de defensa contra virus en plantas. La señal involucrada en el desencadenamiento de la degradación del ARN viral o del especificado por el transgen es producida en la hoja inicialmente infectada (o fotosintéticamente activas en el caso de transgenes), y posteriormente transportada al resto de la planta a través del floema; se ha propuesto que la señal consiste en ácido nucleico, ya sea ADN o ARN (Palauqui et al., 1997). El silenciamiento génico postranscripcional se correlaciona con la presencia de ARNs de pequeño tamaño (entre 21 y 25 nucleótidos) que provienen del ARN silenciado (Hamilton y Baulcombe, 1999). Estos son producidos por la acción de una ARN polimerasa dependiente de ARN y una ribonucleasa específica (Figura 3). Esta última también está involucrada en la generación de micro ARNs que parecen regular diversos genes en plantas y animales (Hutvagner et al., 2001), y se especula que podrían ser las señales móviles que inducen la degradación del ARN blanco. Trabajos más recientes indican que los ARNs de bajo peso molecular son las señales transportadas, si bien ARN de alto peso molecular tampoco puede ser descartado por completo (Klahre et al., 2002).
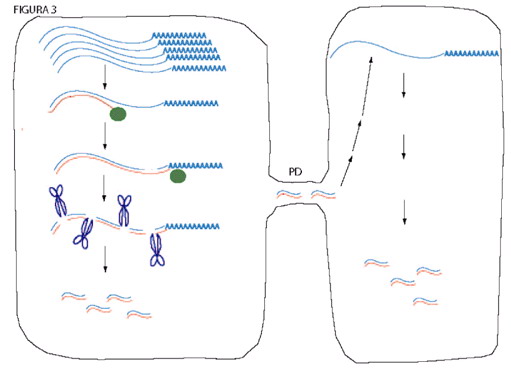
Figura 3. Modelo que representa el silenciamiento génico postranscripcional (PTGS) y su propagación. Un ARN, que puede ser uno endógeno o viral (azul), se acumula a niveles elevados en una célula. Una ARN polimerasa dependiente de ARN (círculo verde) sintetiza ARN complementario a éste (naranja), y una ribonucleasa específica de ARN de cadena doble (representada por unas tijeras) lo corta en fragmentos de 21-25 nucleótidos. Posteriormente, estos fragmentos, de cadena doble o sencilla, o bien, otras moléculas, se desplazan a células vecinas a través de plasmodesmos (PD), en donde inducen la degradación del ARN blanco original.
Dominancia apical
Un fenómeno evidente en una gran variedad de plantas, pero ausente en otras tantas (como arbustos o árboles), es la dominancia apical. Es decir, sólo existe un ápice principal, aun cuando hay meristemos axilares a lo largo del eje de la planta, si bien en estado "latente". Es bien sabido que si el ápice es eliminado, el meristemo axilar más cercano toma su lugar (es decir, se reactivan sus células), ejerciendo su dominio sobre el resto de los meristemos axilares de forma tan "despótica" como el que reemplazó (Napoli et al., 1999). Se desconocen los mecanismos por los que estos meristemos se inactivan y se mantienen latentes. Trabajos recientes han demostrado que se requieren señales provenientes tanto del meristemo apical como de hojas e incluso de la raíz para establecer y mantener la dominancia apical (Booker et al., 2003). Asimismo, se desconoce por completo qué señales están involucradas, aunque se han propuesto que éstas sean auxinas.
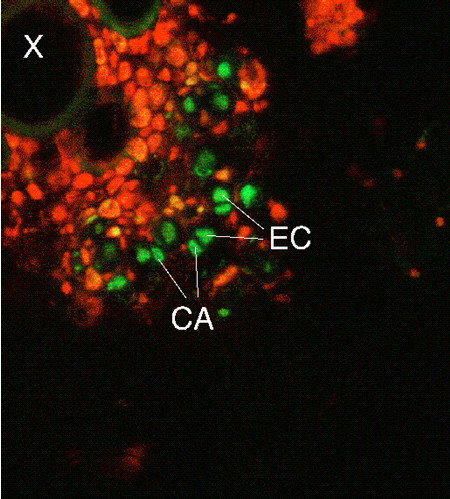
Figura 4. Localización de un ARN mensajero de floema, CmNACP-1, por reacción de polimerización en cadena in situ en un corte transversal de tallo de calabaza (Cucurbita maxima). El color rojo corresponde a la autofluorescencia del tejido por clorofila, y el verde corresponde a la señal producida por el ADN complementario al ARN marcado fluores-centemente. Dicha señal se localiza en el complejo célula acompañante (CA)-elemento criboso (EC). X, vasos del xilema.
Naturaleza de las señales involucradas
La presencia de reguladores del crecimiento en los conductos vasculares de plantas ha sugerido que éstas son señales que comunican tejidos distantes. El papel del ABA como señal que se desplaza entre diferentes tejidos por el tejido vascular (xilema) está bien demonstrado, no así en los casos de las citocininas, etileno, auxinas, ácido giberélico, brasinosteroides u otras moléculas reguladoras del crecimiento vegetal. Su presencia en floema o xilema no es prueba concluyente de que circulen por toda la planta, puesto que, por su tamaño pequeño, podrían difundir de células vecinas (Booker et al., 2003). No obstante, hay evidencia de flujo por el espacio extracelular de reguladores de crecimiento tales como auxinas, etileno, etc., en grupos compactos de células (Taiz & Zeiger, 2002). Los receptores de etileno, auxinas y citocininas son proteínas transmembranales, lo que refuerza la noción de una ruta de transporte extracelular. La presencia de macromoléculas biológicas con propiedades informacionales en el floema sugiere que éstas podrían constituir también señales que intercomunican tejidos distantes. El estudio del transporte de virus dentro de plantas ha apoyado dicha noción.
Transporte de virus y macromoléculas en plantas
Se ha demostrado que diversos virus de plantas se desplazan de una célula a otra a través de los plasmodesmos (Gilbertson et al., 2001) aunque su diámetro útil no permitiría el libre tránsito ni siquiera de los virus más pequeños. Todos los virus de plantas producen una o más de las denominadas proteínas de movimiento, cuya función es aumentar el diámetro útil de los plasmodesmos y transportar los ácidos nucleicos virales. Esto sugirió que proteínas endógenas son capaces de transitar entre células vecinas a través de plasmodesmos. En efecto, ciertas proteínas que regulan el desarrollo de plantas se desplazan de una célula a otra a través de distancias cortas, tanto en el ápice del brote como en la raíz (Lucas et al., 1995; Nakajima et al., 2001). Ya que los virus deben ingresar al floema para infectar de manera sistémica a una planta, se ha propuesto que podrían circular también macromoléculas biológicas endógenas. Por ejemplo, la savia de floema contiene una gran variedad de proteínas capaces de aumentar el límite de exclusión molecular de plasmodesmos (Balachandran et al., 1997). Muchas de estas proteínas están involucradas en el mantenimiento y función de los elementos cribosos. Otras, como la proteína CmPP16 de floema de Cucurbita maxima (calabaza), parecen tener funciones reguladoras. Esta proteína puede unir ARN, independientemente de su secuencia, y transportarlo de una célula a otra a través de plasmodesmos (Xoconostle-Cázares et al., 1999). Esto sugiere que existe ARN mensajero (ARNm) desplazándose en el floema. Efectivamente, la savia de floema contiene una gran variedad de ARNm, algunos de los cuales codifican proteínas que potencialmente regulan el desarrollo de plantas, como CmNACP-1 (Figura 4; Ruiz-Medrano et al., 1999). Desconocemos qué papel puede jugar el transporte en el floema de proteínas o de ARNm. Un mutante de jitomate (mouse eras) sugiere que dicho transporte puede ser relevante en el desarrollo de plantas. La mutación ha sido causada por la fusión de un gen regulador del desarrollo (un gen homeótico) con otro que especifica una enzima involucrada en el metabolismo de azúcares. Experimentos en los que se injertaron plantas silvestres a mutantes demostraron que, sorprendentemente, la apariencia de las hojas surgidas posteriormente en el injerto silvestre era idéntica a la del mutante. Además, la transmisión del fenotipo se debía al transporte del ARN mensajero fusionado del patrón mutante al injerto silvestre a través del floema (Kim et al., 2001). Cabe especular que este fenómeno no es aislado, y que otros ARNs presentes en savia de floema pueden tener funciones reguladoras.
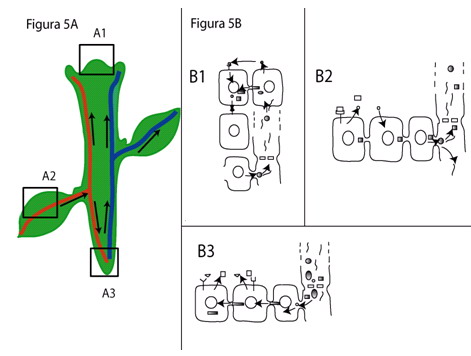
Figura 5. Esquema que ilustra el transporte de señales en diferentes tejidos. (A1, B1) En el meristemo apical ocurre transporte intercelular de proteínas entre las diferentes capas del meristemo
( ![]() ,
,![]() ; pueden ser DEFICIENS y KNOTTED, por ejemplo)
y provenientes del floema, posiblemente de hojas (
; pueden ser DEFICIENS y KNOTTED, por ejemplo)
y provenientes del floema, posiblemente de hojas (![]() ; especulativamente CmPP16).
Este transporte resultaría en la regulación del estado no diferenciado del meristemo apical. Por otra parte, moléculas más pequeñas tales como el péptido
CLAVATA3
; especulativamente CmPP16).
Este transporte resultaría en la regulación del estado no diferenciado del meristemo apical. Por otra parte, moléculas más pequeñas tales como el péptido
CLAVATA3 ![]() se transportan a través del espacio extracelular a células blanco, en donde el receptor correspondiente
CLAVATA1
se transportan a través del espacio extracelular a células blanco, en donde el receptor correspondiente
CLAVATA1 ![]() ,
(ubicado en la membrana plasmática) enlaza estas moléculas. ARNs mensajeros (ARNm) también son transportados al meristemo apical a través del floema,
como CmNACP-1
,
(ubicado en la membrana plasmática) enlaza estas moléculas. ARNs mensajeros (ARNm) también son transportados al meristemo apical a través del floema,
como CmNACP-1 ![]() .
(A2, B2) En una hoja fotosintéticamente activa se ha sugerido que ocurre transporte intercelular de proteínas y ácidos nucleicos vía plasmodesmos. En esta figura se ilustra el caso hipotético de
proteínas (
.
(A2, B2) En una hoja fotosintéticamente activa se ha sugerido que ocurre transporte intercelular de proteínas y ácidos nucleicos vía plasmodesmos. En esta figura se ilustra el caso hipotético de
proteínas (![]() ,
,![]() ) y ARNm
) y ARNm
![]() que son transportados desde el mesófilo, por el floema, al meristemo apical o las raíces. Dentro de dominios de células aledañas ocurre señalización a través del espacio extracelular por medio de fitohormonas como auxinas o
citocininas
que son transportados desde el mesófilo, por el floema, al meristemo apical o las raíces. Dentro de dominios de células aledañas ocurre señalización a través del espacio extracelular por medio de fitohormonas como auxinas o
citocininas ![]() que son enlazadas por receptores
transmembranales
que son enlazadas por receptores
transmembranales ![]() .
Los productos de la fotosíntesis, como
sacarosa
.
Los productos de la fotosíntesis, como
sacarosa ![]() , son transportados tanto por la vía extracelular como intracelularmente. (A3, B3)
En raíces, proteínas como SHR
, son transportados tanto por la vía extracelular como intracelularmente. (A3, B3)
En raíces, proteínas como SHR ![]() se transportan a través de plasmodesmos, mientras que
fitohormonas
se transportan a través de plasmodesmos, mientras que
fitohormonas ![]() son transportadas extracelularmente a células blanco de manera análoga a la descrita en el mesófilo, en donde se unen a receptores
transmembranales
son transportadas extracelularmente a células blanco de manera análoga a la descrita en el mesófilo, en donde se unen a receptores
transmembranales ![]() . Es posible que proteínas
. Es posible que proteínas
![]() y ARNm
y ARNm ![]() , junto con
nutrientes
, junto con
nutrientes ![]() , se desplacen a través del floema de hojas a raíces.
, se desplacen a través del floema de hojas a raíces.
Perspectivas
Como se muestra en la figura 5, es posible que el transporte de señales por la vía extracelular sea más generalizado en el caso de moléculas pequeñas para las que se han encontrado receptores transmembranales, aunque esto no descarta su transporte por el floema. Por otra parte, es probable que el transporte de proteínas y ácidos nucleicos ocurre únicamente a través de plasmodesmos, ya sea entre dominios de células definidos tanto en el meristemo apical del brote (Figura 5; A1, B1), por el floema (A2, B2) como en la raíz (A3, B3), aunque se ignora qué tipo celular se encarga de sintetizarlos. Trabajo adicional en esta dirección determinará cuán general es el transporte y señalización mediada por proteínas y ácidos nucleicos, y su papel en la comunicación intercelular. Desde un punto de vista aplicado, es fácil concebir que el conocer los mecanismos de respuesta a cambios ambientales permitirá la implementación de estrategias racionales de protección de plantas de importancia agrícola contra diversos tipos de estrés ambiental.
Bibliografía
Balachandran S., Xiang, Y., Schobert, C., Thompson, G.A. & Lucas, W.J. (1997). Phloem sap proteins from Cucurbita maxima and Ricinus communis have the capacity to traffic cell to cell through plasmodesmata. Proc. Natl. Acad. Sci. U.S.A., 94: 14150-14155.
Blázquez, M.A., Ahn, J.H. & Weigel, D. (2003). A thermosensory pathway controlling flowering time in Arabidopsis thaliana. Nature Genet., 33: 168-71.
Booker, J., Chatfield, S. & Leyser O. (2003). Auxin acts in xylem-associated or medullary cells to mediate apical dominance. Plant Cell, 15: 495-507.
Colasanti, J. & Sundaresan, V. (2000). 'Florigen' enters the molecular age: long-distance signals that cause plants to flower. Trends Biochem Sci., 25: 236-240
Gilbertson, R.L, Ruiz-Medrano, R., Sudarshana, M. & Lucas, W.J. Plant Virus Movement. In: The Encyclopedia of Plant Pathology. Eds: Malloy, O. C. & Murray, T. D. John Wiley, New York, p.1063.
Hamilton, A.J. & Baulcombe, D.C. (1999). A species of small antisense RNA in posttranscriptional gene silencing in plants. Science, 286: 950-952.
Hutvagner, G., McLachlan, J., Pasquinelli, A.E., Balint, E., Tuschl, T. & Zamore, P.D. (2001). A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science, 293: 834-838.
Jackson, D. (2001). The long and the short of it: signaling development through plasmodesmata. Plant Cell, 13: 2569-2572.
Kim, M., Canio, W., Kessler, S. & Sinha. N. (2001). Developmental changes due to long-distance movement of a homeobox fusion transcript in tomato. Science, 293: 287-289.
Klahre, U., Crete, P., Leuenberger, S.A., Iglesias, V.A. & Meins Jr., F. (2002). High molecular weight RNAs and small interfering RNAs induce systemic posttranscriptional gene silencing in plants. Proc. Natl. Acad. Sci. U.S.A., 99: 11981-11986
Lucas, W.J., Bouche-Pillon, S., Jackson, D.P., Nguyen, L., Baker L., Ding, B. & Hake, S. (1995). Selective trafficking of KNOTTED1 homeodomain protein and its mRNA through plasmodesmata. Science, 270: 1980-1983.
Maldonado, A.M., Doerner, P., Dixon, R.A., Lamb, C.J. & Cameron, R.K. (2002). A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature, 419: 399-403.
Nakajima, K., Sena, G., Nawy, T. & Benfey, P.N. (2001). Intercellular movement of the putative transcription factor SHR in root patterning. Nature, 413: 307-311.
Napoli, C.A, Beveridge, C.A. & Snowden, K.C. (1999). Reevaluating concepts of apical dominance and the control of axillary bud outgrowth. Curr. Top. Dev. Biol., 44: 127-129.
Palauqui, J.-C., Elmayan, T., Pollien, J.M. & Vaucheret, H. (1997). Systemic acquired silencing: transgene-specific post-transcriptional silencing is transmitted by grafting from silenced stocks to non-silenced scions. EMBO J., 16: 4738-4745.
Ruiz-Medrano, R., Xoconostle-Cázares, B. & Lucas, W.J. (1999). Phloem long-distance transport of CmNACP mRNA: implications for supracellular regulation in plants. Development, 126: 4405-4419.
Ruiz-Medrano, R., Xoconostle-Cázares, B. & W.J. Lucas (2001), The phloem as a conduit for inter-organ communication. Curr. Opin. Plant Biol., 4: 202-209.
Ryals, J., Lawton, K.A., Delaney, T.P., Friedrich, L., Kessmann, H., Neuenschwander, U., Uknes, S., Vernooij, B., & Weymann, K. (1995). Signal transduction in systemic acquired resistance. Proc. Natl. Acad. Sci. U S A., 92: 4202-4205.
Ryan, C.A., & Pearce G. (1998). Systemin: a polypeptide signal for plant defensive genes. Annu. Rev. Cell Dev. Biol., 14: 1-17.
Taiz, L. & Zeiger, E. (2002). Plant Physiology (3rd. ed.). Sinauer, New York, NY.
Vance, V. & Vaucheret, H. (2001). RNA silencing in plants--defense and counterdefense. Science 292: 2277-2280.
Xoconostle-Cázares, B., Xiang, Y., Ruiz-Medrano, R., Wang, H.L., Monzer, J., Yoo B.C., McFarland, K.C., Franceschi, V.R. & Lucas, W.J. (1999). Plant paralog to viral movement protein that potentiates transport of mRNA into the phloem. Science, 283: 94-98.
Sitios WWW de interés
Breve enciclopedia de botánica ilustrada
Las definiciones, particularmente aquellas relacionadas con el floema, son bastante actuales, y las figuras poseen una elevada definición. Además es posible vincular con sitios de donde se obtuvo la información original.
http://www.biologie.uni-hamburg.de/b-online/e06/06d.htm
Un tutorial corto ilustrado sobre preguntas frecuentes relacionadas con tejido vascular. Incluye un glosario e ilustraciones y fotografías de gran calidad. http://bugs.bio.usyd.edu.au/2003A+Pmodules/module2.html
Glosario
1. Carbono fijado. Son el conjunto de compuestos orgánicos que sirven de fuente de energía para la planta en los que ha sido incorporado el CO2 atmosférico proveniente de la fotosíntesis.
2. Estoma. Poro microscópico en la epidermis de hoja rodeado por un par de células, denominadas guarda. Los estomas regulan el intercambio de gases en hojas controlando la dimensión del poro.
3. Fitohormonas. Moléculas de diferente naturaleza química que regulan el desarrollo y/o crecimiento de células en plantas. Generalmente son de bajo peso molecular (menos de 1 kD) y en muchos casos sus actividades se traslapan. Por otra parte, estas moléculas parecen ser sintetizadas por los mismos tipos celulares que responden a ellas, y, a diferencia de las hormonas en animales, la gran mayoría de células de una planta son capaces de sintetizarlas.
4. Meristemo. Conjunto de células proliferativas sin identidad determinada y que origina el cuerpo de la planta. Se encuentran en diferentes partes de la planta, tales como en el ápice del brote y de la raíz, tejido vascular, etc.
5. MicroARN. ARNs de pequeño tamaño (menos de 50 nucleótidos) que se ha propuesto regulan la expresión de genes con quienes comparten parte de su secuencia. Se especula que la regulación ocurre por un mecanismo semejante al silenciamiento génico postranscripcional; es decir, por la formación de ARN de cadena doble, que induce la degradación del ARN blanco.
6. Placa cribosa. Pared que delimita elementos cribosos vecinos que contiene poros.
7. Plasmodesmos. Canales membranosos que interconectan el citoplasma de células vecinas a través de la pared celular en plantas. Consisten en una porción de retículo endoplásmico apresado en medio de dicho canal, y permiten el movimiento selectivo de moléculas por la acción tanto de proteínas virales como de planta.
8. Primordio. Grupo de células ya comprometidas a diferenciarse en un órgano determinado, por ejemplo, hoja o raiz, proveniente de un meristemo.
9. Regulacion transcripcional. La transcripción es el proceso por el cual se sintetiza ARN a partir de ADN como molde. El ARN producido puede ser traducido a proteína (como es el caso del ARN mensajero para un gen determinado) o no (como los ARNs ribosómicos o de transferencia). La gran mayoría de los genes de un organismo no se transcriben continuamente; en general ARNs específicos son sintetizados únicamente en respuesta a estímulos o en tejidos determinados. Es decir, la regulación transcripcional es el conjunto de mecanismos que controlan la producción de ARN de un gen en respuesta a estos estímulos o en un tejido dado en organismo multicelular.
10. Vecinos anticlinales. Son grupos de células que tienen un linaje común y cuyo plano de división es perpendicular al eje principal de la planta.