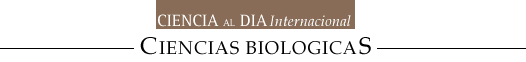
MEDICINA |
Patogénesis de la nefropatía diabética |
|
© M.
Carmen Iglesias-de la Cruz, Sheldon Chen,
Fuad N. Ziyadeh y Miguel A. Pancorbo-Alonso, 2003 mccruz@iib.uam.es |
RESUMEN[ABSTRACT]
En la patogénesis de la nefropatía diabética
son importantes los factores genéticos, hemodinámicos y metabólicos.
Diferentes mediadores y vías de transducción de señales
interactúan en una serie de mecanismos autocrinos, paracrinos y endocrinos
cuando el riñón es expuesto crónicamente a un ambiente
con alta concentración de glucosa. El efecto de esta alta concentración
de glucosa en las células renales puede manifestarse a través
de un incremento de su metabolismo hacia la vía del poliol, incremento
de la síntesis de novo de diacilglicerol con activación
de la proteín kinasa C, activación de la vía de la hexosamina
o aumento de la glicación no enzimática de proteínas. El
aumento de presión sanguínea, primero dentro de los capilares
glomerulares y después sistémicamente, y las consecuencias metabólicas
de los niveles anormalmente altos de glucosa en las células renales,
son los elementos del concepto más ampliamente aceptado de cómo
se desarrolla y progresa la nefropatía diabética. Ambas anormalidades
inducen una respuesta maladaptativa en la que se sobreproduce Factor de Crecimiento
Transformante-b
(según sus siglas en inglés TGF-b), lo que estimula una síntesis
excesiva de moléculas de matriz extracelular. La expansión del
mesangio glomerular y la fibrosis intersticial caracterizan patológicamente
a la nefropatía diabética, y son el resultado de una acumulación
de exceso de matriz.
Antes del descubrimiento de la insulina y su utilización clínica como tratamiento de fondo de la diabetes, el coma diabético era la complicación más frecuente y a menudo fatal, lo que condicionaba una esperanza de vida muy limitada para el enfermo. Con la utilización de la insulina esto dejó de ser así; los diabéticos dejaron de estar condenados a morir jóvenes a causa de un coma y, por tanto, se prolongó notablemente la supervivencia. No obstante, el prolongar la vida de los pacientes ha traído como consecuencia la aparición de complicaciones desconocidas en el pasado, que continúan siendo desafíos casi tres cuartos de siglo más tarde.
La nefropatía diabética, una complicación grave de la diabetes, es una de esas complicaciones debidas a que los pacientes diabéticos viven más tiempo en la actualidad. La enfermedad renal afecta a un tercio de los pacientes diabéticos, y la diabetes es la causa más habitual de fracaso renal en los países desarrollados (Breyer, 1992). A pesar de las mejoras en el control de los niveles de glucosa en sangre y en el manejo de la hipertensión, pocos avances se han producido hacia la curación o la prevención de la nefropatía diabética. Conforme la función renal decae, los pacientes diabéticos se enfrentan sucesivamente a la anemia, la acidosis crónica, el desequilibrio electrolítico, la enfermedad metabólica ósea, la hipertensión, el síndrome nefrótico y la uremia (Mogyorosi & Ziyadeh, 1996). Los pacientes con nefropatía diabética también presentan un riesgo extremadamente alto de enfermedad cardiovascular, con una tasa de mortalidad 30 veces superior a la población general (Mogyorosi & Ziyadeh, 1996). A pesar de los avances en diálisis y en transplante renal, los pacientes diabéticos con enfermedad renal terminal aun tienen una tasa de mortalidad más alta que los pacientes no diabéticos que sufren similares alteraciones renales.
El coste económico de la nefropatía diabética continúa en alza. Conforme los pacientes se aproximan a un estado terminal de la enfermedad renal, son necesarias grandes cantidades de dinero para los cuidados domiciliarios del paciente, la medicación, las visitas a urgencias y las repetidas hospitalizaciones. Una vez que alcanzan la enfermedad terminal, afrontar el transplante renal supone un gasto importante.
El riñón tiene como principal función el filtrado de la sangre que le llega por la arteria renal. Cada riñón tiene en su corteza alrededor de un millón de minúsculas unidades llamadas nefronas, y es en ellas donde se lleva a cabo la filtración. Las nefronas están formadas por una pequeña bolsa llamada cápsula de Bowman. En el interior de esta bolsa se agrupa un entramado de capilares sanguíneos, que en su conjunto se denomina glomérulo. En los vasos del glomérulo se produce la filtración y el producto de esta filtración es depositado en el espacio de la cápsula de Bowman; éste se comunica con unos conductos tubulares, donde tras sufrir diversos procesos de concentración, llega a los uréteres. El marcador clínico más temprano de la enfermedad renal diabética es la microalbuminuria, definida como presencia en la orina de 20 a 200 microgramos por minuto ó 30 a 300 miligramos por 24 horas de la proteína más abundante en sangre, la albúmina. Atacar el problema de la nefropapía diabética requiere un entendimiento más profundo del proceso de esta enfermedad.
Este artículo se centra en los mecanismos patogénicos de la nefropatía diabética, con énfasis en los factores hemodinámicos y metabólicos. El conocimiento actual está aún lejos de ser completo y sin duda motivará futuras investigaciones.
Cambios estructurales que ocurren precozmente en la diabetes
La estructura del riñón está profundamente alterada en prácticamente todos los pacientes diabéticos, incluso en aquellos no destinados a desarrollar nefropatía diabética establecida, la cual se define como albuminuria mayor de 300 miligramos por día y progresiva disminución en el filtrado glomerular. Uno de los primeros cambios, que ocurre a los pocos meses del diagnóstico de la diabetes, es el engrosamiento renal causado por la hipertrofia celular dentro de los túbulos y glomérulos. Casi todos los pacientes desarrollarán engrosamiento de las membranas basales glomerular y tubular a los 2 ó 3 años del diagnóstico de la diabetes. Sin embargo, sólo algunos llegarán a desarrollar expansión del mesangio glomerular y fibrosis intersticial, las marcas patológicas de la nefropatía diabética progresiva.
El primer signo clínico fiable de una nefropatía incipiente es la aparición de microalbuminuria. Con el tiempo, la nefropatía evoluciona y clínicamente se manifiesta como la aparición de proteinuria, hipertensión e insuficiencia renal. Existe una buena correlación entre la expansión de la región del mesangio, la severidad de la fibrosis intersticial y la arteriosclerosis, con el descenso de la tasa de filtración glomerular (TFG) (Ziyadeh & Goldfarb, 1991). Ello tiene como consecuencia que la expansión mesangial reduce la filtración glomerular por oclusión de los capilares glomerulares y disminución del área efectiva para la filtración. De la misma manera, la fibrosis tubulointersticial altera la arquitectura y función tubular y ello conduce a insuficiencia renal (Ziyadeh & Goldfarb, 1991). Debido a la importancia de la expansión del mesangio y la fibrosis tubulointersticial, la investigación se ha centrado en los mecanismos que conducen a la acumulación de matriz, tanto al aumento en la síntesis como la disminución en su degradación.
TGF-beta como principal agente etiológico
En la última década se ha propuesto a una molécula efectora como el principal agente etiológico de la nefropatía diabética. El factor de crecimiento transformante b (según sus siglas en inglés, TGF-beta) es una citoquina hipertrófica que media virtualmente en todos los cambios patológicos de la nefropatía diabética. Al igual que un ambiente alto en glucosa, el TGF-beta causa hipertrofia en las células renales y promueve la producción de un exceso de matriz extracelular en las células glomerulares, tubulares y en los fibroblastos intersticiales. Niveles altos de glucosa estimulan aún más la producción de TGF-beta y la síntesis de sus receptores en los compartimentos glomerular y tubulointersticial (Hoffman et al., 1998; Rocco et al., 1992). En presencia de concentraciones normales de glucosa, el TGF-beta administrado exógenamente produce el mismo daño renal que la hiperglicemia.
La manipulación genética para aumentar la expresión del TGF-beta en los glomérulos de ratas normales conduce a glomerulosclerosis. Los ratones transgénicos que sobreproducen TGF-beta desarrollan proteinuria, glomerulosclerosis y fibrosis tubulointersticial. Finalmente, la inhibición de la actividad del TGF-beta previene en gran medida los efectos dañinos de la alta glucosa en células mesangiales en cultivo, reduciendo la hipertrofia y previniendo la acumulación de matriz (Wolf et al., 1992). El bloqueo de la bioactividad del TGF-beta también ralentiza la progresión de la insuficiencia renal en ratones diabéticos (Sharma et al., 1996; Ziyadeh et al., 2000).
Mucho se ha aprendido en los últimos años acerca de la fisiología de la acción del TGF-beta. El TGF-beta causa hipertrofia renal interfiriendo en la regulación del ciclo celular. En presencia del TGF-beta, una célula se detendrá en la fase G1 del ciclo celular, un periodo caracterizado por síntesis de proteínas con ausencia de replicación de ADN, lo cual es la definición de hipertrofia. El TGF-beta por tanto difiere de otras citoquinas en que éstas últimas generalmente provocan hiperplasia. El TGF-beta también promueve la acumulación de matriz extracelular a través de la estimulación de su síntesis y la inhibición de su degradación. Este último efecto se consigue disminuyendo la actividad de las enzimas que digieren la matriz (colagenasa y diversas metaloproteinasas) o aumentando la actividad de los inhibidores de dichas enzimas (inhibidores del activador del plasminógeno). A través de estos mecanismos el TGF-beta puede mostrar un potente efecto profibrótico (Sharma & Ziyadeh, 1994). Su función es importante en la reparación normal de los tejidos, pero una actividad excesiva causa una producción de matriz exuberante que lleva a una fibrosis patológica, lo cual podría amplificar la respuesta fibrótica de una manera retroalimentativa.
Influencia de los cambios de presión arterial en la respuesta adaptativa renal
Una de las grandes teorías acerca de la susceptibilidad a la enfermedad renal afirma que tanto la hipertensión arterial sistémica como una presión aumentada en los capilares glomerulares, son precursores de la glomerulosclerosis diabética (Mogensen, 1985). La elevada presión puede dañar las células endoteliales, epiteliales y mesangiales; así pues juega un papel significativo en la iniciación y progresión del proceso de esclerosis.
En pacientes con diabetes tipo 1, y en menor medida en la diabetes tipo 2, se observa una TFG aumentada en un 25-50%, debido principalmente a incrementos en el flujo plasmático renal y a la presión capilar glomerular. Algunos datos de ensayos clínicos sugieren que tener una elevada TFG pone al paciente diabético en un gran riesgo de desarrollar nefropatía. Un estudio de ocho años en pacientes con diabetes tipo 1 mostró que el 53% de ellos con una TFG inicial de 125 ml por minuto, desarrolló microalbuminuria o nefropatía manifiesta, frente a sólo un 5% entre los que tenía una TFG menor (Dahlquist et al., 2001). Estudios clásicos de micropunción glomerular realizados por el Dr. Barry M. Brenner y colaboradores, demostraron en un modelo de nefrona aislada, que la rata moderadamente hiperglicémica tiene una elevada TFG (Hostetter et al., 1981). Este aumento en la TFG parece estar mediado por un incremento en el flujo sanguíneo y la presión capilar. Además, como parte de la hipertrofia en los estadios iniciales de la enfermedad diabética, el área capilar glomerular se agranda, contribuyendo al incremento de la TFG (Hostetter et al., 1981). Hay que tener en cuenta la dinámica situación que afecta a la TFG. Ésta es elevada durante el inicio de la diabetes y se mantiene de este modo mientras la excreción de albúmina se mantenga normal o persista la microalbuminuria. Una vez que se desarrolla la macroalbuminuria, la TFG desciende progresivamente (Nelson et al., 1996).
El tono vascular en las arteriolas aferente y eferente es también un determinante mayor del flujo y presión capilar glomerular. En la diabetes, la arteriola aferente se relaja, permitiendo un mayor flujo sanguíneo al glomérulo, mientras que la arteriola eferente permanece relativamente comprimida e incrementa la presión hidrostática en el glomérulo. Cuando la hipertensión sistémica aparece en un paciente diabético, la alta presión arterial es transmitida al glomérulo, elevando más aún la presión hidrostática. Esta hipertensión intraglomerular crónica lleva a la expansión de la matriz mesangial, pérdida de área de filtración y descenso progresivo en la TFG. Finalmente, la expansión del diámetro del capilar glomerular que ocurre precozmente en el curso de la diabetes, incrementa proporcionalmente la tensión de las paredes capilares, de acuerdo con la ley de Laplace: la tensión es proporcional a la presión transmural y al radio del vaso.
Recientemente se ha establecido una intrigante asociación hemodinámica-metabólica en la evolución de la nefropatía diabética. El estiramiento cíclico de células mesangiales in vitro, lo cual mimetiza la hipertensión intraglomerular, estimula marcadamente la actividad del TGF-beta y causa aumento en la síntesis de colágeno y otros componentes de matriz. Más aún, un ambiente alto en glucosa actúa sinérgicamente con el estiramiento cíclico para inducir al TGF-beta (Riser et al., 1998). La activación del TGF-beta renal parece ser un mediador común de virtualmente todos los factores que están implicados en el desarrollo y progresión de la nefropatía diabética.
Efectos directos e indirectos de la hiperglicemia
La hiperglicemia es el sine qua non de la diabetes mellitus y se ha sospechado hace tiempo que puede ser la causa de las complicaciones diabéticas. Sorprendentemente la conexión no fue fácil de mostrar, pero dos estudios clínicos han probado definitivamente que la alta concentración de glucosa es fundamental para el desarrollo de la nefropatía. El Diabetes Control and Complications Trial demostró esta asociación para la diabetes tipo 1 en 1993; el United Kingdom Prospective Diabetes Study hizo lo mismo para la diabetes tipo 2 en 1998. Más recientemente, otro estudio reseñable mostró que un riguroso control de la hiperglicemia a largo plazo (10 años), conseguido por transplante pancreático, pudo incluso revertir las lesiones patológicas de nefropatías establecidas en pacientes con diabetes tipo 1 (Fioretto et al., 1998). La pregunta que queda por contestar es por qué los altos niveles de glucosa son tóxicos para el riñón. Con técnicas cada vez más sofisticadas, esa pregunta está empezando a ser contestada a un nivel molecular y celular. Los efectos de la glucosa elevada sobre células renales pueden aparecer debido a un incremento en el metabolismo de la glucosa a través de la vía del poliol, una síntesis de novo incrementada de diacilglicerol con la consecuente activación de proteín kinasas y de la vía de las hexosaminas, o por un incremento en la glicación no enzimática de proteínas. La siguiente exposición da un repaso a las consecuencias metabólicas de la alta glicemia, activación resultante de vías celulares y alteración de la fisiología que culmina con la enfermedad renal diabética.
La elevada concentración de glucosa en el espacio extracelular define el estado diabético, pero la tasa del metabolismo de la glucosa intracelular dicta el desarrollo de la nefropatía diabética. Las células renales no requieren de insulina para la captación de la glucosa, pues poseen transportadores transmembrana que mueven la glucosa a través de la membrana celular. La glucosa difunde debido a un gradiente menor de concentración, y la captación, la fosforilación y el metabolismo de la glucosa incrementan en proporción a la concentración de glucosa circulante.
Entre las distintas isoformas de transportadores de glucosa (GLUT), la más ampliamente expresada en el riñón es GLUT-1. Una peculiaridad del riñón en el estado diabético es que un ambiente alto en glucosa regula positivamente la transcripción génica y la producción proteica de GLUT-1 en células mesangiales. Esto provoca mayor captación de glucosa y su utilización celular. Se ha mostrado que sobrexpresando GLUT-1 en células mesangiales de rata la captación de glucosa se incrementó incluso cuando las células fueron cultivadas con concentraciones normales de glucosa, las células se comportaron como si estuvieran cultivadas en alta glucosa, manifestando incremento de síntesis de proteínas de matriz (Heilig et al., 1995). Por lo tanto, es el flujo incrementado de glucosa a través de múltiples vías metabólicas intracelulares el responsable de la acumulación de matriz extracelular en células mesangiales.
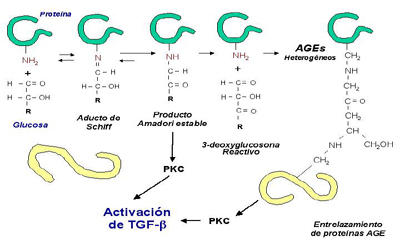
Figura 1. Metabolismo de la glucosa
Las células del riñón utilizan glucosa en varias rutas de su metabolismo. El metabolismo de la glucosa a través de la glucolisis (Fig 1) y la vía del poliol favorece la formación de diacilglicerol (DAG), el activador endógeno primario de la proteín kinasa C que está implicadas en la secreción de factores de crecimiento y síntesis de matriz extracelular. La glucolisis desempeña el doble papel de degradar la glucosa para generar ATP y provee sustancias básicas para otras rutas anabólicas. A medio camino de la glucolisis, la glucosa se disocia en dos azúcares de tres carbonos, el gliceraldehido-3-fosfato (GAP) y la dihidroxiacetona fostato (DHAP). La DHAP puede ser convertida en GAP, el cual continuará hacia los siguientes pasos de la glicolisis, o puede ser reducida a glicerol-3-fosfato, el mayor precursor de la síntesis de diacilglicerol. El flujo aumentado de glucosa a través de la vía del poliol ayuda a generar el poder reductor que es necesario para convertir el DHAP a glicerol-3-fosfato. La glucosa es convertida primero a sorbitol y después oxidada a fructosa, con reducción de NAD+ a NADH. La acumulación en tejidos de sorbitol per se, puede jugar un cierto papel en la formación de las cataratas en la diabetes a través de un efecto osmótico, pero otros efectos indirectos de la vía del poliol pueden ser operativos en otras complicaciones diabéticas como la nefropatía. Por ejemplo, el cociente intracelular NADH/ NAD+ incrementado crea un estado redox favorable para la conversión de DHAP a GAP. Cuando el GAP es abundante, la célula puede sintetizar diacilglicerol, el cual teóricamente puede activar las cascadas de señales dependientes de la proteín kinasa C (Ziyadeh, 1995).
El papel de la vía del poliol (Fig 1) en el desarrollo de la nefropatía diabética aún es controvertido. Los resultados de la inhibición de la aldosa reductasa, la enzima limitante en la vía del poliol, han sido ineficaces en la prevención o el tratamiento de la nefropatía diabética experimental. Una razón para dichos resultados puede ser la incompleta inhibición de la enzima por los agentes disponibles en la actualidad (Daniels & Hostetter, 1989).
Alguno de los efectos reguladores de la glucosa elevada pueden ser mediados por otra vía metabólica, la vía de las hexosaminas, en la cual la fructosa-6-fosfato (F6P) se convierte en glucosamina-6-fosfato, que a continuación pierde su grupo amido para convertirse en glutamato (Fig 1). Los productos finales de esta vía son uridina difosfato-N-acetil glucosamina y otras hexosaminas nucleotídicas. La aminación de la F6P es el paso limitante y es catalizada por la glutamina-F6P amidotransferasa (GFAT). Un estudio reciente ha sugerido que la producción de TGF-beta inducida por la alta glucosa esta mediada por la vía de la hexosamina (Kolm-Litty et al., 1998). Este estudio ha mostrado que la D-glucosamina, un análogo estructural de la D-glucosa, aumenta la producción del TGF-beta y la fibronectina en células mesangiales, y que la inhibición de la GFAT con oligonucleótidos antisentido previene el incremento en la expresión del TGF-beta inducido por la alta glucosa (Kolm-Litty et al., 1998).
La hiperglicemia también puede tener efectos adversos sobre el riñón a través de la formación de proteínas glicadas. La glicación es un proceso no enzimático en el cual un azúcar reductor, como la glucosa, se une de manera covalente a una proteína circulante (Fig 2). Esta modificación de la proteína puede alterar su función, promover estrés oxidativo intracelular, y conducir a la activación de vías metabólicas importantes en la patogénesis de la enfermedad renal diabética. El primer producto estable de la glicación se forma en un período de días a semanas. La glucosa reacciona con el grupo amino de la proteína circulante para formar un aducto llamado de Schiff. Este producto intermedio se reorganiza pasando a ser un producto Amadori. El segundo mayor grupo de proteínas glicadas son los productos de glicosilación avanzada o AGEs y derivan de una serie de reordenamientos, deshidrataciones y condensaciones de los productos Amadori (Fig 2). Los AGEs se forman en períodos de meses y se mantienen durante toda la vida de la proteína. La evidencia de que los productos Amadori son capaces de dañar al riñón han venido fundamentalmente de trabajos experimentales. En nuestro laboratorio hemos cultivado células mesangiales glomerulares y endoteliales en medio de cultivo que contenía albúmina modificada Amadori, la cual es la principal proteína glicada en el suero. Comparado con la albúmina normal, la albúmina glicada estimula la expresión génica del TGF-beta y las moléculas de matriz extracelular como el colágeno tipo IV y la fibronectina. Un anticuerpo monoclonal específico contra la albúmina glicada previene estos cambios (Cohen et al., 1994).
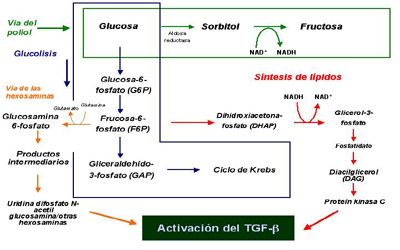
Figura 2. Glicación de proteinas.
Los estudios in vivo con albúmina modificada Amadori en ratones diabéticos han sido igualmente rotundos. El tratamiento de ratones db/db (modelo murino de diabetes tipo 2) durante ocho semanas con el anticuerpo anti-albúmina glicada disminuye las concentraciones de albúmina glicada en suero, reduce la proteinuria, detiene la expansión mesangial y previene la expresión incrementada de moléculas de matriz extracelular en la corteza renal (Cohen et al., 1995). Una de las consecuencias cruciales del incremento de la albúmina glicada circulando por el riñón es la activación de sistemas de señalización intracelular, sobre todo la PKC, y la consiguiente estimulación del sistema del TGF-beta (Chen et al., 2001).
Las evidencias experimentales disponibles también demuestran la participación de los productos de glicosilación avanzada en la progresión de la nefropatía diabética. Con el desarrollo de técnicas que detectan AGEs en muestras de tejido, numerosos grupos han mostrado que los AGEs se acumulan en los riñones de humanos diabéticos. Cuando se inyecta un AGE (albúmina modificada en forma de AGE) a ratones normales durante ocho semanas desarrollan hipertrofia glomerular asociada con incremento en el ARNm del TGF-beta (Cohen et al., 2002). Un primer paso hacia el entendimiento sobre cómo los AGEs exhiben sus efectos dañinos ha sido el receptor para los AGEs (RAGE) recientemente descubierto. Aunque los eventos intracelulares causados por la activación de este receptor RAGE aun no han sido totalmente caracterizados, observaciones empíricas sugieren una interacción AGE-RAGE que causarían efectos como la producción de fibronectina o de colágeno tipo IV. Anticuerpos para los RAGE bloquean este efecto, indicando que la respuesta es dependiente del receptor de AGE.
La aminoguanidina, un inhibidor de la formación de AGEs, se ha mostrado prometedora en el tratamiento de la nefropatía diabética (Ellis & Good, 1991). Puede atenuar la albuminuria en diferentes modelos animales de diabetes y puede prevenir la expansión mesangial y el engrosamiento de las membranas basales glomerulares (Soulis-Liparota et al., 1991). Debido a estos hechos, se han desarrollado ensayos clínicos en humanos usando este agente, en los cuales la proteinuria se redujo en numerosos pacientes, pero los resultados en general fueron decepcionantes debido a la considerable toxicidad.
Los radicales libres de oxígeno se producen de manera excesiva en diversos
estados patológicos y concretamente pueden contribuir a la patogénesis
de diferentes enfermedades renales agudas o crónicas que cursan con glomerulosclerosis
y fibrosis tubulointersticial, como la nefropatía diabética (Trachtman
et al., 1995). Diversos estudios han demostrado que los radicales libres de
oxígeno producen un aumento de diversas moléculas que intervienen
en síntesis de matriz extracelular, proliferación y contracción
celular en el riñón (Scharschmidt & Dunn, 1983; Trachtman
et al., 1995). Por último, se ha demostrado que estos radicales libres
de oxígeno producen un aumento en la expresión y actividad de
TGF-beta y como consecuencia se incrementa la producción de proteínas
de matriz extracelular en células mesangiales en cultivo (Iglesias-de
la Cruz et al., 2001). Estas respuestas celulares al estrés oxidativo
fueron inhibidas en presencia de un anticuerpo anti-TGF-beta, demostrando así
que un desequilibrio en la producción de moléculas oxidantes pueden
jugar un papel determinante en el desarrollo y progresión de enfermedades
renales caracterizadas por un exceso de matriz extracelular, como es el caso
de la nefropatía diabética (Iglesias-de la Cruz et al., 2001).
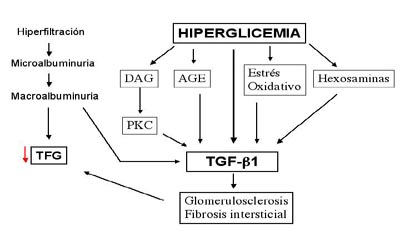
Figura 3. Papel del TGF-beta y de la hiperglicemia, en la nefropatía diabética
En la actualidad se cree que la alta proteinuria per se, es un factor independiente en la progresión de todas las formas de enfermedad crónica renal. La excesiva filtración de proteínas dentro del túbulo proximal y su consecuente endocitosis puede representar un mecanismo patológico importante subyacente a la cicatrización renal. Un posible escenario para explicar la relación entre el grado de proteinuria y la insuficiencia renal progresiva puede ser la reabsorción tubular de proteínas y la infiltración de macrófagos. Los macrófagos, además de ser una fuente rica en TGF-beta, promueven la transformación de fibroblastos inactivos a miofibroblastos activados. La síntesis y depósito por parte de estas células de colágenos fibrosos tipos I y III, contribuye a la aparición de fibrosis tubulointersticial, cuya severidad se correlaciona con el descenso de la TFG. Se establece un círculo vicioso según el cual la pérdida de nefronas aumenta el estrés hemodinámico glomerular en las nefronas supervivientes, aumenta la proteinuria y acelera el daño tubulointersticial. Por lo tanto, fármacos que produzcan una disminución en la proteinuria y en la expresión renal del TGF-beta romperían el mencionado círculo vicioso.
La figura 3 representa un esquema que bien podría resumir el papel de algunos de los mediadores, con un papel central del TGF-beta y la hiperglicemia, de la nefropatía diabética y su situación cronológica en función del desarrollo de la enfermedad, en relación también con los cambios morfológicos y funcionales que se aprecian.
Resumiendo, el ideal sería la propia prevención de la diabetes mellitus, pero este objetivo hoy por hoy es inalcanzable. El abordaje terapéutico de la enfermedad renal diabética se ha centrado en el control de la glicemia y de la presión arterial. Pero estas maniobras simplemente han retrasado, mas que evitado, la aparición del estado terminal de la enfermedad renal. En un futuro próximo el tratamiento de la diabetes y sus complicaciones debe involucrar un enfoque multidisciplinario que debería incluir transplante pancreático, manipulaciones genéticas para eliminar la predisposición hereditaria y maniobras específicas para interceptar el eje del TGF-beta que eviten los efectos dañinos de la fibrosis.
Referencias
Breyer, J. A. (1992). Diabetic nephropathy in insulin-dependent
patients. Amer. J of Kidney Dis, 20 (6), 533-547.
Chen, S., Cohen, M. P., Lautenslager, G. T., Shearman, C.
W., & Ziyadeh, F. N. (2001). Glycated albumin stimulates TGF-beta1 production
and protein kinase C activity in glomerular endothelial cells. Kidney Int, 59:
673-681.
Cohen, M. P., Sharma, K., Jin, Y., Hud, E., Wu, V. Y., Tomaszewski,
J., & Ziyadeh, F. N. (1995). Prevention of diabetic nephropathy in db/db
mice with glycated albumin antagonists. A novel treatment strategy. J Clin Invest,
95: 2338-2345.
Cohen, M. P., Sharma, K., Yulin, J., & Ziyadeh, F. N.
(1994). Treatment with monoclonal antibody to glycated albumin prevents diabetic
nephropathy in db/db mice. Clin. Res., 42: 143A.
Cohen, M. P., Ziyadeh, F. N., Hong, S. W., Shearman, C. W.,
Hud, E., Lautenslager, G. T., Iglesias-de la Cruz, M. C., & Chen, S. (2002).
Inhibiting albumin glycation in vivo ameliorates glomerular overexpression of
TGF-beta eta1. Kidney Int, 61: 2025-2032.
Dahlquist, G., Stattin, E. L., & Rudberg, S. (2001).
Urinary albumin excretion rate and glomerular filtration rate in the prediction
of diabetic nephropathy; a long-term follow-up study of childhood onset type-1
diabetic patients. Nephrol Dial Transplant, 16: 1382-1386.
Daniels, B., & Hostetter, T. (1989). Aldose reductase
inhibition and glomerular abnormalities in diabetic rats. Diabetes, 38: 981-986.
Ellis, E. N., & Good, B. H. (1991). Prevention of glomerular
basement membrane thickening by aminoguanidine in experimental diabetes mellitus.
Metabolism, 40: 1016-1019.
Fioretto, P., Steffes, M. W., Sutherland, D. E., Goetz, F.
C., & Mauer, M. (1998). Reversal of lesions of diabetic nephropathy after
pancreas transplantation. N Engl J Med, 339: 69-75.
Heilig, C. W., Concepcion, L. A., Riser, B. L., Freytag,
S. O., Zhu, M., & Cortes, P. (1995). Overexpression of glucose transporters
in rat mesangial cells cultured in a normal glucose milieu mimics the diabetic
phenotype. J Clin Invest, 96: 1802-1814.
Hoffman, B. B., Sharma, K., Zhu, Y., & Ziyadeh, F. N.
(1998). Transcriptional activation of transforming growth factor-beta1 in mesangial
cell culture by high glucose concentration. Kidney Int., 54: 1107-1116.
Hostetter, T. H., Troy, J. L., & Brenner, B. M. (1981).
Glomerular hemodynamics in experimental diabetes mellitus. Kidney Int, 19: 410-415.
Iglesias-de la Cruz, M. C., Ruiz-Torres, M. P., Alcamí,
J., Díez-Marqués, M. L., Ortega-Velázquez, R., Chen, S.,
Rodríguez-Puyol, M., Ziyadeh, F. N., & Rodríguez-Puyol, D.
(2001). Hydrogen peroxide increases extracellular matrix mRNA through TGF-beta
in human mesangial cells. Kidney Int, 59: 87-95.
Kolm-Litty, S. U., Nerlich, A., Lehmann, R. & Schleider,
E.D. (1998). High glucose-induced transforming growth factor beta1 production
is mediated by the hexosamine pathway in porcine glomerular mesangial cells.
J Clin Invest, 101: 160-169.
Mogensen, C. E. (1985). Longterm antihypertensive treatment
inhibiting progression of diabetic nephropathy. Br Med J., 285: 685-688.
Mogyorosi, A., & Ziyadeh, F. N. (1996). Update on pathogenesis,
markers and management of diabetic nephropathy. Curr Opin Nephrol Hypertens,
5: 243-253.
Nelson, R. G., Bennett, P. H., Beck, G. J., Tan, M., Knowler,
W. C., Mitch, W. E., Hirschman, G. H., & Myers, B. D. (1996). Development
and progression of renal disease in Pima Indians with non-insulin-dependent
diabetes mellitus. Diabetic Renal Disease Study Group. N Engl J Med, 335: 1636-1642.
Riser, B. L., Cortes, P., Yee, J., Sharba, A. K., Asano,
K., Rodriguez-Barbero, A., & Narins, R. G. (1998). Mechanical strain- and
high glucose-induced alterations in mesangial cell collagen metabolism: role
of TGF-beta. J Am Soc Nephrol, 9: 827-836.
Rocco, M. V., Chen, Y., Goldfarb, S., & Ziyadeh, F. N.
(1992). Elevated glucose stimulates TGF-beta gene expression and bioactivity in
proximal tubule. Kidney Int, 41: 107-114.
Scharschmidt, L. A., & Dunn, M. J. (1983). Prostaglandin
synthesis by rat glomerular mesangial cells in culture. Effects of angiotensin
II and arginine vasopressin. J Clin Invest, 71: 1756-1764.
Sharma, K., Jin, Y., Guo, J., & Ziyadeh, F. N. (1996).
Neutralization of TGF-beta by anti-TGF-beta antibody attenuates kidney hypertrophy
and the enhanced extracellular matrix gene expression in STZ-induced diabetic
mice. Diabetes, 45: 522-530.
Sharma, K., & Ziyadeh, F. N. (1994). The emerging role
of transforming growth factor-beta in
Soulis-Liparota, T., Cooper, M., Papaoglou, D., Clarke, B.,
& Jerums, G. (1991). Retardation by aminoguanidine of development of albuminuria,
mesangial expansion and tissue fluorescence in streptozocin-induced diabetic
rat. Diabetes, 40: 1328-1334.
Trachtman, H., Futterweit, S., Maesaka, J., Ma, C., Valderrama,
E., Fuchs, A., Tarectecan, A. A., Rao, P. S., Sturman, J. A., Boles, T. H.,
Fu, M.-X., & Baynes, J.(1995). Taurine ameliorates chronic streptozocin-induced
diabetic nephropathy in rats. Am J Physiol, 269: F429-F438.
Trachtman, H., Futterweit, S., & Singhal, P. (1995).
Nitric oxide modulates the synthesis of extracellular matrix proteins in cultured
rat mesangial cells. Biochem Biophys Res Commun, 207: 120-125.
Wolf, G., Sharma, K., Chen, Y., Ericksen, M., & Ziyadeh,
F. N. (1992). High glucose-induced proliferation in mesangial cells is reversed
by autocrine TGF-beta. Kidney Int, 42: 647-656.
Ziyadeh, F. N. (1995). Mediators of hyperglycemia and the
pathogenesis of matrix accumulation in diabetic renal disease. Miner Electrolyte
Metab, 21: 292-302.
Ziyadeh, F. N., & Goldfarb, S. (1991). The renal tubulointerstitium
in diabetes mellitus. Kidney Int, 39: 464-475.
Ziyadeh, F. N., Hoffman, B. B., Han, D. C., Iglesias-de la Cruz, M. C., Hong, S. W., Isono, M., Chen, S., McGowan, T. A., & Sharma, K. (2000). Long-term prevention of renal insufficiency, excess matrix gene expression and glomerular mesangial matrix expression by treatment with monoclonal anti-TGF-beta antibody in db/db diabetic mice. Proc. Natl. Acad. Sci. USA, 97: 8015-8020.
Sitios de Interes
Enciclopedia Médica en español
http://www.nlm.nih.gov/medlineplus/spanish/ency/article/000494.htm
Geosalud, un sitio medico en la WWW
http://geosalud.com/diabetesmellitus/diabetesnefropatia.htm
Sitio sobre Nefrología (en inglés)
http://www.nephrologychannel.com/diabeticnephropathy/
Sitio sobre salud y patologías de la Universidad de
Pennsylvania (en ingés)
http://www.hmc.psu.edu/healthinfo/d/diabeticnephropathy.htm