
BIOLOGIA |
Bases moleculares de la floración |
RESUMEN [ABSTRACT]
El éxito en la reproducción de las plantas depende de la floración sincronizada de todos los individuos de una misma población y de la correcta construcción de los órganos de la flor, encontrándose ambos procesos bajo control ambiental y genético. Utilizando una pequeña planta, Arabidopsis thaliana, hemos podido alcanzar en los últimos años un conocimiento relativamente satisfactorio acerca de los mecanismos moleculares por los que las plantas reconocen las condiciones ambientales y modifican la expresión génica con el objetivo último de construir las flores. El conjunto de instrucciones parece estar conservado en muchas otras especies vegetales, lo que va a permitir, en un futuro no tan lejano, manipular la floración a voluntad en especies más útiles que Arabidopsis.
Tanto los biólogos aficionados como los poetas han gozado desde hace miles de años con la enorme variedad de formas, tamaños, y colores con que nos regalan las flores. Sin embargo, sólo hemos comenzado a entender cómo se construyen éstas hace relativamente poco tiempo. Hay tres cuestiones básicas que las plantas deben solucionar para reproducirse con éxito: cuándo florecer, dónde han de brotar las flores, y cómo construir los órganos florales. Hoy en día conocemos relativamente bien cuáles son los mecanismos moleculares por los que las plantas reconocen las condiciones ambientales, transducen la señal al interior y finalmente establecen el programa de desarrollo floral. Este avance en el conocimiento se debe sobre todo al éxito del abordaje genético en Arabidopsis thaliana, una mala hierba adoptada como modelo vegetal desde los años 80 (Figura 1).

Figura 1. Arabidopsis thaliana. A la izquierda se muestra una planta en la fase vegetativa de crecimiento, en la que sólo se producen hojas. A la derecha se muestra una planta en la fase reproductora, en la que la planta produce flores en lugar de hojas, alarga el tallo principal y genera tallos secundarios en las axilas de hojas previamente formadas.
Dicha estrategia ha consistido en el aislamiento de mutantes afectados en cada uno de las etapas de floración (Figura 2): unos con el tiempo de floración alterado (más tardíos o tempranos que las plantas silvestres); otros con defectos en la iniciación de las flores individuales; y algunos con defectos en la identidad de los organos florales (por ejemplo, con pétalos sustituyendo a estambres, etc.) El análisis del fenotipo de todas estas plantas mutantes y de las interacciones entre ellas, así como la identificación de los genes afectados en cada una de ellas ha permitido no sólo conocer nuevos procesos desde el punto de vista básico, sino también comenzar a aplicar dichos conocimientos para manipular la floración a voluntad, como veremos más adelante.
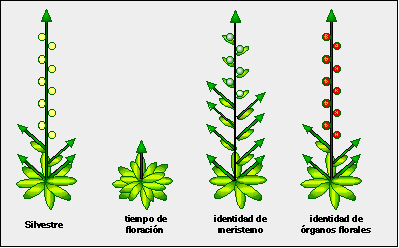
Figura 2. Mutantes de floración en Arabidopsis.
Tiempo de floración
Arabidopsis es, desde el punto de vista del tiempo de floración, una planta facultativa de día largo. Esto significa que Arabidopsis puede florecer en un régimen de días cortos (unas 8-10 h de luz al día, equivalente al otoño/invierno), pero los días largos (unas 16 h de luz al día, equivalente a primavera/verano) promueven una floración mucho más temprana (tres semanas desde la germinación de las semillas, frente a dos meses en días cortos).
Entre los numerosos mutantes afectados en el tiempo de floración se pueden distinguir al menos dos grupos: aquellos que florecen más tarde sólo en días largos, y aquellos que florecen más tarde tanto en días largos como en días cortos. Esto indica que los primeros definen una vía de señalización que se activa preferentemente en días largos, mientras que los segundos participan en la promoción de la floración independientemente de cuál sea el fotoperíodo. El análisis más refinado de las interacciones genéticas entre los distintos mutantes y el aislamiento de los genes afectados en cada caso ha permitido establecer una jerarquía entre ellos -es decir, qué genes actúan antes o después en la transducción de las señales de floración- así como definir las distintasvías de señalización. El escenario que emerge de todos estos resultados es una complicada trama de vías de señalización (Figura 3) activadas cada una de ella por una condición (ambiental o endógena) particular, pero con conexiones entre ellas (Koornneef et al., 1998; Levy & Dean, 1998).
Una pregunta esencial, aún no respondida en su totalidad, es cómo se integra la información generada por estas señales. Caben dos posibles escenarios: o bien cada vía actúa independientemente sobre los genes que inician el desarrollo floral (es decir, la información se integraría en las dianas últimas), o bien hay elementos comunes a varias de estas vías (que actuarían, de este modo, como integradores). Existen candidatos a actuar como integradores de la vía que no son los genes de iniciación del desarrollo floral. Por ejemplo, el mutante emf1 (embryonic flower 1) florece después de apenas formar dos hojas, cuando la plántula acaba de germinar. Por esta razón se ha propuesto que la floración sería un programa de desarrollo que se activaría por defecto, pero reprimido en condiciones normales por EMF1. Según esta hipótesis, la promoción de la floración por las vías antes mencionadas se efectuaría mediante la progresiva supresión de la actividad del represor EMF1. Esta hipótesis, aunque atractiva, carece aún de evidencia experimental. Al contrario, lo que sí se ha podido demostrar, como veremos más adelante, es que son los promotores de los genes diana los que actuarían como integradores.
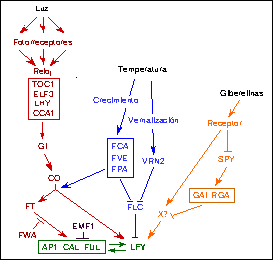
Figura 3. Esquema de las vías de inducción floral y sus dianas moleculares en Arabidopsis. Consultar el Glosario de Términos para más información sobre "vías de señalización".
Iniciación floral
Las flores de Arabidopsis se inician en las regiones laterales alrededor del meristemo apical del tallo, y nacen de los mismos primordios que dan lugar a hojas durante la fase vegetativa de crecimiento. Que estos primordios den lugar a una flor en lugar de formar una hoja depende de la actividad de proteínas como LEAFY (LFY). A modo de interruptor, la presencia de LFY por encima de cierto nivel umbral dispara el proceso de desarrollo floral, mientras que su ausencia impide la formación de flores (Weigel et al., 1992). La expresión de LFY está regulada por todas las vías que controlan el tiempo de floración. Por ejemplo, el defecto de floración del mutantega1 en días cortos está causado por una falta de activación de la expresión de LFY, mientras que la expresión forzada de LFY a partir de un promotor constitutivo en el mutante ga1 suprime el defecto de floración.
En el promotor de LFY se ha identificado la secuencia a través de la cual las giberelinas regulan la expresión de este gen. La eliminación de esta secuencia ha permitido, por ejemplo, construir plantas que florezcan exclusivamente en días largos, pero no en días cortos, convirtiendo a Arabidopsis en una planta obligatoria -en lugar de facultativa- de días largos. Este resultado indica, además, que el promotor de LFY actúa directamente como integrador de al menos dos vías de promoción de la floración: la de días largos y la dependiente de giberelinas (Blázquez & Weigel, 2000).
LFY no es el único gen de identidad de meristemo floral. La proteína codificada por APETALA1 (AP1) y otros genes relacionados estructuralmente (CAULIFLOWER y FRUITFULL) también son necesarias para conferir a un primordio la identidad floral (Ferrándiz et al., 2000; Liljegren et al., 1999). La expresión de AP1 parece estar controlada, al igual que la de LFY, por una rama de la vía facultativa de días largos. Sin embargo, LFY juega el papel más importante en este proceso puesto que su expresión precede a la de AP1 y el fenotipo del mutante lfy es más grave que el de ap1. Por otra parte, el que las plantas de Arabidopsis continúen formando flores en los flancos del ápice hasta que envejecen, en lugar de consumir el ápice en la formación de una flor terminal -como sucede en otras especies- depende de la actividad de proteínas como TFL1, que impiden que LFY y AP1 extiendan su dominio de expresión al centro del ápice.
¿De qué forma activa LFY el desarrollo inicial de la flor? LFY es un factor de transcripción necesario para la expresión de los genes maestro que dividen el primordio floral en las regiones que van a dar lugar a los órganos de la flor adulta. En los promotores de estos genes se encuentran secuencias reconocidas por LFY y a través de las cuales induce su expresión. En esta labor, LFY no actúa solo. De lo contrario sería complicado explicar cómo LFY, que está presente a lo largo y ancho de todo el primordio floral, puede provocar la activación de genes en sólo ciertas regiones del primordio. Sin embargo, aún no se ha identificado ningún coactivador de LFY.
Identidad de órganos florales
El programa de desarrollo floral consite en la expresión regional de ciertos genes, similares en función a los genes homeóticos de animales, que instruyen a las células en la formación de un determinado órgano. El primordio floral es, a simple vista, un grupo homogéneo de células, sin que se distingan diferencias morfológicas. Sin embargo, muy pronto en el desarrollo existen diferencias a nivel de expresión génica, en el interior de las células. El primordio floral puede dividirse en cuatro regiones concéntricas, denominadas verticilos, que coinciden con las células que darán lugar a sépalos, pétalos, estambres y pistilos (Figura 4).

Figura 4. Flor típica de Arabidopsis.
La combinación del análisis genético de mutantes deficientes en la formación de determinados órganos, con el análisis molecular de los genes afectados en cada mutante, permitió proponer el ya clásico "modelo ABC" del desarrollo floral (Coen & Meyerowitz, 1991). Según este modelo (Figura 5), los genes de tipo A (AP1, APETALA2) se expresan en los dos verticilos externos, los de tipo C (AGAMOUS) en los dos interiores, y los de tipo B (APETALA3, PISTILLATA) se expresan en los verticilos 2 y 3. La actividad de los genes A establece la identidad de los sépalos en el primer verticilo y, junto con la actividad B, la de pétalos en el segundo verticilo. De forma análoga, B y C juntos especifican identidad de estambres en el tercer verticilo, mientras que la actividad de C establece por sí misma la identidad del pistilo. También se han identificado genes (SEPALLATA1,2, y 3) que se expresan en los tres verticilos interiores, y presentan actividad B y C. La expresión de A impide la de C y viceversa. Por esta razón, en un mutante sin actividad de tipo A, por ejemplo, la expresión de genes de tipo C se extiende a todos los verticilos externos, y las flores de este mutante contienen la secuencia de órganos: pistilo, estambres, estambres, pistilo.
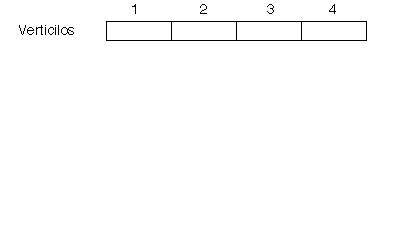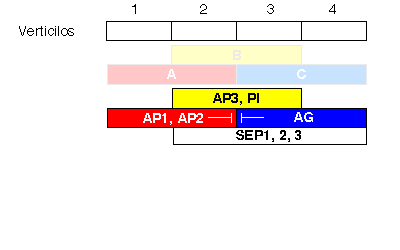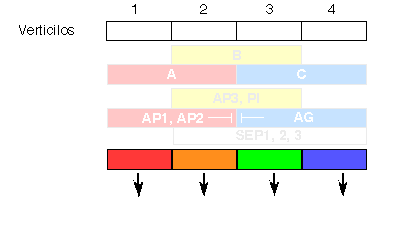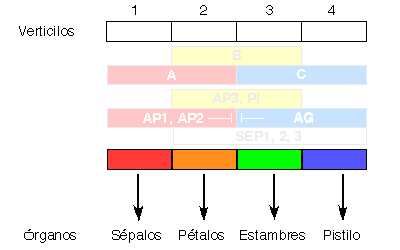
Figura 5. Modelo "ABC" para la determinación de la identidad de los órganos florales.
Los genes de identidad de desarrollo floral, excepto AP2, codifican proteínas del tipo "MADS box". Estas proteínas son factores de transcripción, por lo que es fácil imaginar que los estadíos posteriores de la construcción de cada uno de los órganos de la flor requerirá la activación secuencial o coordinada de ciertos genes, y que esta regulación dependerá de los genes ABC. Aunque hay dianas formales por debajo de la acción de los genes de identidad de órganos (enzimas para la síntesis de pigmentos en pétalos, etc), aún no se han identificado las dianas primarias a cuyos promotores se unan las mencionadas proteínas de tipo "MADS box". Por esta misma razón, es difícil comprender cómo la identidad de ciertos órganos la establece exclusivamente un tipo de proteínas (por ejemplo, AP1 para sépalos), mientras que la de otros órganos requiere la acción combinada de dos o más tipos de proteínas (por ejemplo, AP1, AP3 y PI para pétalos). Las proteínas de tipo "MADS box" pueden formar, en el tubo de ensayo, dímeros con ellas mismas o con otros miembros de la familia, pero para saber si ésta es la manera en que funcionan en la planta es necesario identificar en primer lugar alguna diana funcional para estos factores de transcripción.
Consideraciones finales
Si uno de los objetivos de la investigación en este campo es ser capaz de manipular el desarrollo reproductor (tanto sus aspectos temporales como la arquitectura de las flores), una pregunta relevante es hasta qué punto puede extrapolarse a otras especies de mayor interés aplicado lo aprendido sobre la floración de Arabidopsis. Las perspectivas son buenas. Por una parte, los programas de secuenciación de genomas de plantas están identificando continuamente genes que son similares estructuralmente a los genes que controlan la floración en Arabidopsis. Por ejemplo, se conocen ortólogos de LFY o AP1 en tabaco, arroz, guisante, maíz, tomate, y árboles como el pino y el álamo.

Figura 6. Floración temprana de álamos inducida por la expresión forzada del gen LEAFY de Arabidopsis bajo el control del promotor constitutivo viral 35S. Las flechas indican la ausencia de flores en el álamo silvestre, y la presencia de flores en el álamo transgénico (Imagen cortesía de D. Weigel y O. Nilsson).
Este hallazgo, por sí mismo, no implica que la floración esté controlada precisamente por dichos genes en especies distintas a Arabidopsis; para comprobarlo será necesario realizar un análisis funcional de los mismos, y esto no es siempre posible en algunas de las especies citadas. Pero dos resultados resultan esperanzadores: por una parte, los genes LFY de tabaco, pino y maíz pueden sustituir al gen LFY en Arabidopsis; y, de forma análoga, la expresión constitutiva y elevada del gen LFY de Arabidopsis en álamo provoca la floración temprana (Weigel & Nilsson, 1995) (Figura 6). Además, se han encontrado variedades mutantes de trigo o arroz que florecen más tarde y se encuentran probablemente afectadas en un gen similar a CO, lo cual, de confirmarse, indicaría que el programa de floración en Arabidopsis y otras plantas se realiza con guiones y actores similares, incluso si estas otras plantas presentan hábitos de floración tan diversos (independientes del fotoperíodo, o dependientes de días cortos, etc.)
Referencias bibliográficas
Blázquez, M.A. & Weigel, D. (2000) Integration of floral inductive signals in Arabidopsis. Nature 404: 889-892.
Coen, E.S. & Meyerowitz, E.M. (1991) The war of the whorls: genetic interactions controlling flower development. Nature 353: 31-37.
Ferrándiz, C., Gu, Q., Martienssen, R. & Yanofsky, M.F. (2000) FRUITFULL involvement in floral meristem identity in Arabidopsis. Development 127: 725-734.
Koornneef, M., Alonso-Blanco, C., Peeters, A.J.M. & Soppe, W. (1998) Genetic control of flowering in Arabidopsis Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 345-370
Levy, Y.Y. & Dean, C. (1998) The transition to flowering. Plant Cell 10, 1973-1990.
Liljegren, S.J., Gustafson-Brown, C., Pinyopich, A. & Yanofsky, M.F. (1999) Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 specify meristem fate. Plant Cell 11: 1007-1018.
Weigel, D. & Nilsson, O. (1995) A developmental switch sufficient for the initiation of flowers in diverse species. Nature 377: 495-500.
Weigel, D., Alvarez, J., Smyth, D.R., Yanofsky, M.F. & Meyerowitz, E. M. (1992) LEAFY controls floral meristem identity in Arabidopsis. Cell 69: 843-859.
Punteros de Interés
TAIR: The Arabidopsis Information Resource (www.arabidopsis.org). El punto de encuentro oficial para todos los investigadores de Arabidopsis. Bases de datos, herramientas útiles, información general.
The Flowering Web (www.salk.edu/LABS/pbio-w/flower_web.html). Información actualizada periódicamente sobre los genes que controlan el tiempo de floración en Arabidopsis.
Weigel Lab Website (www.salk.edu/LABS/pbio-w). Información general sobre regulación de la floración, y sobre LEAFY en particular.
The Meyerowitz Lab Website.(www.caltech.edu/~meyerowitz/). Colección de imágenes de mutantes afectados en la identidad de órganos florales e información general sobre el desarrollo floral.
Yanofsky Lab Website (www-biology.ucsd.edu:80/others/yanofsky/home.html). Abundante información sobre genes de tipo "MADS-box" implicados en el desarrollo floral.
Dean Lab Website (www.jic.bbsrc.ac.uk/staff/caroline-dean/index.htm). Información general sobre inducción floral, y sobre el papel de la vernalización en particular.
