
BIOLOGIA |
El Transporte Intracelular de Proteínas: El caso del TGF-alfa. |
|
© Jesús M. Ureña y Joaquín Arribas, 2000
jarribas@hg.vhebron.es, Unidad de Investigación Oncológica del Hospital Valle de Hebron de Barcelona |
RESUMEN [ABSTRACT]
Las diferentes proteínas celulares han de ser clasificadas y transportadas a lugares concretos de la célula donde llevarán a cabo su función. En el caso de las proteínas de membrana, una vez sintetizadas en los ribosomas son translocadas al Retículo Endoplasmático (RE) gracias a la existencia de unas "secuencias señal" que las dirigen a este destino. En el RE sufren modificaciones como glucosilación y proteólisis y son transportadas al Aparato de Golgi (AG) donde sufren nuevas modificaciones y son finalmente clasificadas para su destino final que puede ser la secreción extracelular o la incorporación a la membrana plasmática. Estudiando este proceso para una molécula de transmembrana de la superficie celular: el Factor de Crecimiento Transformante Alfa (TGF-alfa), hemos encontrado que una señal necesaria para su correcto transporte se encuentra en el último aminoácido de su secuencia peptídica, que es una Valina. Una mutación en este aminoácido provoca la retención de la molécula en el RE, interrumpiéndose su transporte. Investigando la maquinaria de transporte implicada en esta señal, hemos encontrado una molécula que participa en el proceso, se encuentra en el RE y forma una interacción estable con TGF-alfa silvestre, pero no con mutantes en la Valina carboxi-terminal. Esta proteína recibe el nombre de TACIP18 y, probablemente, es el primer componente de una vía molecular de transporte de proteínas con motivos intracelulares similares
Las células animales se caracterizan por llevar a cabo un gran número de funciones diferentes a través de proteínas cuya actuación, además de estar coordinada, ha de tener lugar en un lugar concreto y específico. Así, ciertas proteínas, tales como receptores de hormonas, de factores de crecimiento o de adhesión han de participar en funciones que tienen lugar en la membrana plasmática. En el núcleo ejercen su función moléculas del metabolismo del ácido desoxirribonucleico como polimerasas y nucleasas. Otras proteínas se localizan en las mitocondrias (enzimas sintetizadores de ATP), en los lisosomas (proteasas y otras enzimas degradativas), o en el citosol (enzimas de la glucólisis, capaces de obtener energía a partir de la combustión de azúcares). Otras proteínas han de ser secretadas fuera de la célula, como en el caso de las células de glándulas exocrinas que exportan hormonas al torrente circulatorio. Finalmente, otras proteínas han de ser almacenadas en unos gránulos secretores donde permanecen hasta que llegue una señal de secreción que desencadenará el proceso de liberación de estas moléculas (neurotransmisores, transportadores de glucosa) a la membrana celular o al exterior. Para clasificar cada proteína, dirigirla a su destino correcto y controlar el momento correcto de su activación, la célula utiliza mecanismos que estamos empezando a conocer y que ilustran la complejidad de los procesos biológicos.
Una vez que una proteína es sintetizada en los ribosomas puede permanecer en la parte soluble del citoplasma (citosol), ser dirigida a mitocondrias o el núcleo, puede ser enviada a la membrana plasmática o celular, o a organelos internos como lisosomas. ¿Cómo se consigue este proceso de clasificación y reparto? En 1970, Sabatini y Blobel demostraron que existía una señal, que denominaron péptido señal en la zona amino-terminal (la primera en ser sintetizada) de ciertas proteínas que hacía que se dirigieran al Retículo Endoplasmático (RE). El RE es una estructura membranosa presente en las células eucariotas que se encuentra rodeando al núcleo celular. En el RE se produce una interacción entre el péptido señal de la proteína naciente y receptores específicos que provoca la translocación al interior del RE de la nueva proteína en un proceso dependiente de la energía de hidrólisis del ATP (Blobel & Dobberstein, 1975; Sudhoff, 1995). En el lumen del RE, las proteínas son desprovistas del péptido señal y sufren modificaciones tales como adición de azúcares (glucosilación). Después las proteínas son dirigidas al Aparato de Golgi (AG) donde sufren nuevas modificaciones y finalmente son dirigidas bien a los lisosomas, a vesículas secretoras o a la membrana plasmática. El envío de proteínas desde los ribosomas libres al núcleo o las mitocondrias sigue otra vía diferente en la que participan señales presentes en la secuencia primaria de aminoácidos de las proteínas como las "secuencias de localización nuclear" (NLS)
El péptido señal que dirige las proteínas a la vía RE-AG-membrana/secreción/lisosomas varía entre 13 y 36 aminoácidos, y es eliminado en el lumen del RE por una peptidasa. Otras modificaciones tienen lugar en este compartimento, se forman los puentes disulfuro de las proteínas (entre aminoácidos cisteina) que contribuyen a que las proteínas se plieguen adquiriendo una conformación funcional, y muchas proteínas son glucosiladas. La glucosilación consiste en la adición de oligosacáridos unidos a asparagina por puentes N-glucosídicos (N-glucosilación) o a serina y treonina por puentes O-glucosídicos (O-glucosilación) en secuencias de aminoácidos concretas. Algunos antibióticos, como la Tunicamicina o la Bacitracina, son capaces precisamente de bloquear estos pasos. Las proteínas presentes en el lumen del RE y en la membrana del RE son transportadas al Aparato de Golgi, un conglomerado de sacos membranosos que tiene dos funciones principales: la modificación y elaboración de unidades de carbohidratos y la clasificación de las moléculas para el siguiente destino que serán los lisosomas, la membrana plasmática o gránulos de secreción. El Aparato de Golgi a su vez se diferencia en tres compartimentos: Cis-Golgi (proximal al RE), Golgi Medio y Trans-Golgi donde realmente se produce la exportación de las proteínas (Rothman, 1994).
El RE es un orgánulo membranoso compuesto por una red de cisternas. Las proteínas residentes en el RE se clasifican en cinco grupos: proteínas implicadas en la translocación y el transporte vesicular, enzimas implicados en el plegamiento de proteínas, enzimas implicados en modificaciones post-traduccionales, enzimas implicados en biosíntesis de lípidos, y enzimas implicados en el mantenimiento y almacenamiento de Ca2+. En tipos celulares concretos encontramos enzimas específicas como el Citocromo 450 en hepatocitos que juega un papel de detoxificación. El transporte de las proteínas desde el RE al Cis-Golgi tiene lugar mediante la formación de unas vesículas de transporte que se producen por gemación desde el RE y se fusionan con la membrana del Cis-Golgi en un proceso en el que participan proteínas evolutivamente conservadas desde la levadura al ser humano. Existen dos tipos de vesículas que se denominan COP1 y COPII con funciones diferenciadas (Teasdale & Jackson, 1996). Los carbohidratos añadidos en el RE son modificados en cada uno de los compartimentos del AG en un proceso denominado glucosilación terminal frente al proceso de glucosilación central que tiene lugar en el RE. Este proceso de glucosilación terminal contendría la O-glucosilación y modificaciones de N-glucosilación. Se asume que el RE es una estación de control de calidad que impide que las proteínas incorrectamente plegadas (y por tanto no funcionales), progresen a través de la vía secretora. ¿Como se consigue que las proteínas residentes del RE se mantengan en esta estructura y no sean conducidas por la vía secretora a la membrana? Pelham y colaboradores en 1989 demostraron la existencia de una secuencia peptídica presente en el extremo carboxílico de ciertas proteínas residentes en el RE que era necesaria y suficiente para retener las proteínas en el RE, observaron que cuando esta secuencia era incorporada a una enzima secretada (la lisozima, un agente antibacteriano), la enzima híbrida permanecía retenida en el RE. Esta secuencia está formada por los aminoácidso Lys-Asp-Glu-Leu (KDEL según la simbología de una letra utilizada para designar aminoácidos). Posteriormente se han descrito otras secuencias que también son señales de retención en el RE: señales lys-lys-X-X y señales X-X-Arg-Arg (donde X puede ser cualquier aminoácido). Por otro lado, Nishimura y Balch (1997) han demostrado que una secuencia concreta (Asp-X-Asp) en la glicoproteína del virus de la estomatitis vesicular contiene la información necesaria para el paso de esta proteína desde el RE al AG. Cuando esta señal se muta, dicha proteína queda retenida y no progresa en su transporte.
En la actualidad se cree que existen dos mecanismos distintos para asegurar el mantenimiento de proteínas en el RE: la retención por exclusión activa y el retrotransporte desde el AG al RE. Esta segunda posibilidad ha sido demostrada tanto para el motivo KDEL como para el motivo di-lisina (lys-lys-X-X). También se ha visto que otras proteínas jamás abandonan el orgánulo, y se supone que estas proteínas no se llegan a incorporar a las vesículas que se generan en el RE. Uno de los modelos propuestos para explicar este fenómeno sería a través de la oligomerización.
Una vez en el AG, algunas proteínas son enviadas a lisosomas. Conocemos bien este proceso gracias al estudio que se ha hecho de una enfermedad denominada Mucolipidosis tipo II o enfermedad I-celular (Amara et al., 1992). En esta enfermedad los pacientes presentan un severo retraso psicomotor, así como deformidades esqueléticas, sus lisosomas presentas inclusiones de carbohidratos (de ahí el nombre I) y glucolípidos no digeridos. Estas inclusiones intracelulares se deben a la ausencia de 8 hidrolasas necesarias para la degradación de carbohidratos en lisosomas. La ausencia de estas hidrolasas en los lisosomas se correlaciona con su aumento muy importante en plasma y orina, lo que hizo pensar que el defecto se encontraba en el transporte de estas enzimas que, en lugar de ser conducidos a los lisosomas, eran llevados al exterior de la membrana plasmática por la vía secretora. El análisis bioquímico de estas hidrolasas permitió comprobar que las enzimas de estos pacientes no contenían el azúcar Manosa-6-fosfato, que sí se encuentra en las enzimas de las personas no afectadas. Este azúcar se encuentra unido a la proteína. La proteína adquiere la Manosa-6-fosfato en el compartimento Cis-Golgi gracias a la actuación de un enzima fosfotransferasa que es precisamente el causante de la enfermedad. Así pues, ahora sabemos que las proteínas que son dirigidas a los lisosomas sufren la adición de una molécula de Manosa 6-fosfato. Esta "etiqueta" dirige estas moléculas hasta los lisosomas donde existe un receptor de Manosa 6-fosfato con el que interaccionan las proteínas lisosomales y a continuación se produce la translocación al lisosoma de esta molécula.
Las proteínas que no son dirigidas a lisosomas desde el AG, serán secretadas de manera constitutiva o regulada. Se empaquetarán en vesículas de secreción que al fusionarse con la membrana plasmática o celular, bien por defecto o en respuesta a señales específicas, expondrán las proteínas del lumen vesicular al espacio extracelular. Ciertas proteínas con segmentos hidrofóbicos permitirán el anclaje a los lípidos de la membrana y quedarán como proteínas integrales de membrana que podrán sufrir procesos de maduración subsiguientes; otras serán secretadas y difundirán al espacio extracelular.
El transporte a la superficie celular es un proceso menos conocido. Se presume el transporte por defecto. Nosotros hemos estudiado el papel que juega una señal presente en el extremo carboxi-terminal de proteínas como el TGF-alfa y las Metaloproteasas de matriz de tipo transmembrana (MT-MMP) en el transporte y la localización subcelular de estas proteínas. Hemos visto que una señal tan simple como un solo aminoácido valina en el extremo C-terminal (intracelular) juega un papel determinante en cuanto al tráfico correcto de esta molécula a lo largo de todo el trayecto subcelular que comprende desde la síntesis en los ribosomas a su localización en la membrana.
El TGF-alfa: Transporte intracelular y maduración
El TGF-alfa es un factor de crecimiento que se une a un receptor de superficie celular llamado "Receptor del Factor de Crecimiento Epidérmico" (EGFR), activándolo y promoviendo en la célula que contiene el receptor fenómenos de división celular. El TGF-alfa es sintetizado como una proteína integral de membrana denominada proTGF-alfa que contiene un dominio de transmembrana y una corta cola en el citoplasma. El dominio extracelular puede ser liberado generando una señal mitogénica que actúa a una distancia considerable de la célula que lo ha producido. El sistema de liberación de TGF-alfa puede ser activado por una kinasa intracelular, la Proteína Kinasa C (PKC, Bosemberg et al., 1992; Pandiella & Massagué, 1991), y parece ser un sistema celular general que actúa sobre distintas moléculas de superficie que pueden sufrir esta clase de corte o proteólisis (Arribas et al., 1996). De forma alternativa, una vez que el factor de crecimiento se encuentra anclado en la membrana también puede activar el EGFR en las células adyacentes. En la membrana plasmática tiene lugar un segundo fenómeno de maduración consistente en la proteolización de un fragmento N-terminal (extracelular) altamente glucosilado que deja finalmente la proteína madura y activa (Arribas et al., 1997). La porción citoplasmática del TGF-alfa contiene 34 aminoácidos, es rica en cisteinas (hay 7 en total) y está muy conservada entre especies, lo que sugiere un importante papel para esta zona de la proteína. Inicialmente se había descrito que esta zona era importante para la liberación del fragmento extracelular de TGF-alfa mediado por PKC (Bosemberg et al., 1992), siendo crítico en este proceso el último aminoácido en el extremo C-terminal (valina). Mediante experimentos en los que se mutan aminoácidos en el extremo C-terminal hemos visto que esta parte de la proteína juega un papel necesario para la maduración y el transporte a la superficie celular, habiendo determinado que el determinante concreto es el último aminoácido valina (Ureña et al., 1999; Briley et al., 1997). El mecanismo de biosíntesis del TGF-alfa puede ser saturado mediante sobre-expresión transitoria de proteína normal, pero no es saturado por la sobrexpresión de proteína con mutaciones en el extremo C-terminal o con proteínas cuyo transporte a la superficie no es dependiente de la porción citoplasmática, como por ejemplo en el caso del beta-glicano.
Como hemos discutido, la mayoría de proteínas son clasificadas de acuerdo con señales contenidas en su secuencia. Para determinar la posible existencia de una señal que dirigiera a pro-TGF-alfa a la superficie celular, realizamos una serie de proteínas mutantes tal y como se muestra en la Figura 1. Una de estas proteínas mutantes, presentaba el extremo intracelular modificado (aunque poseía el resto de la proteína intacto) y no era detectable en la superficie de la célula tal y como puede verse en la figura 2A. En el panel superior se presenta la localización de membrana de la proteína silvestre, frente al nivel de detección en superficie disminuido de la proteína mutada en el panel inferior. Únicamente detectamos la proteína en la superficie si la porción citoplasmática estaba intacta. A continuación determinamos las alteraciones mínimas de la porción citoplasmática que interferían con el transporte de pro-TGF-alfa y pudimos comprobar que la simple mutación de un único aminoácido en el extremo carboxi-terminal, provoca una drástica reducción de los niveles detectados de proteína de superficie, indicando que este aminoácido (valina) juega un papel sustancial en el proceso. El transporte a la membrana plasmática de otras proteínas como el beta-glicano, no se veía afectado por la delección o mutación de su zona intracelular y su detección era indistinguible entre la proteína normal y la mutada en la superficie celular.

Figura 1. Obtención de moléculas mutantes de pro-TGF-alfa. El esquema muestra la estructura de la proteína silvestre del pro-TGF-alfa. El recuadro en negro muestra la zona correspondiente al péptido señal, en sombreado el dominio de TGF-alfa, y la región de rayas corresponde al dominio de transmembrana. El dominio intracelular corresponde a los aminoácidos 125-159. Se realizaron mutantes en los que se había eliminado 31 aminoácidos (pro-HA-TGF-alfa D31), 3 aminoácidos (pro-HA-TGF-alfa DC4) y mutaciones en las que el último aminoácido valina se cambiaba por glicina ( proHA-TGF-alfa V159G), serina (proHA-TGF-alfa V159S) o ácido glutámico (proHA-TGF-alfa V159E).
Los mutantes de TGF-alfa, en los cuales la valina del extremo citoplasmático había sido sustituida por cualquier otro aminoácido (ver Figura 2), se localizan en el RE, indicando que no son transportados eficientemente a la superficie, como puede observarse en forma de fluorescencia intermedia (color naranja) en esta estructura (compárese con el marcaje de una proteína residente en el RE: proteína disulfido isomerasa: PDI) (Fig. 2B).
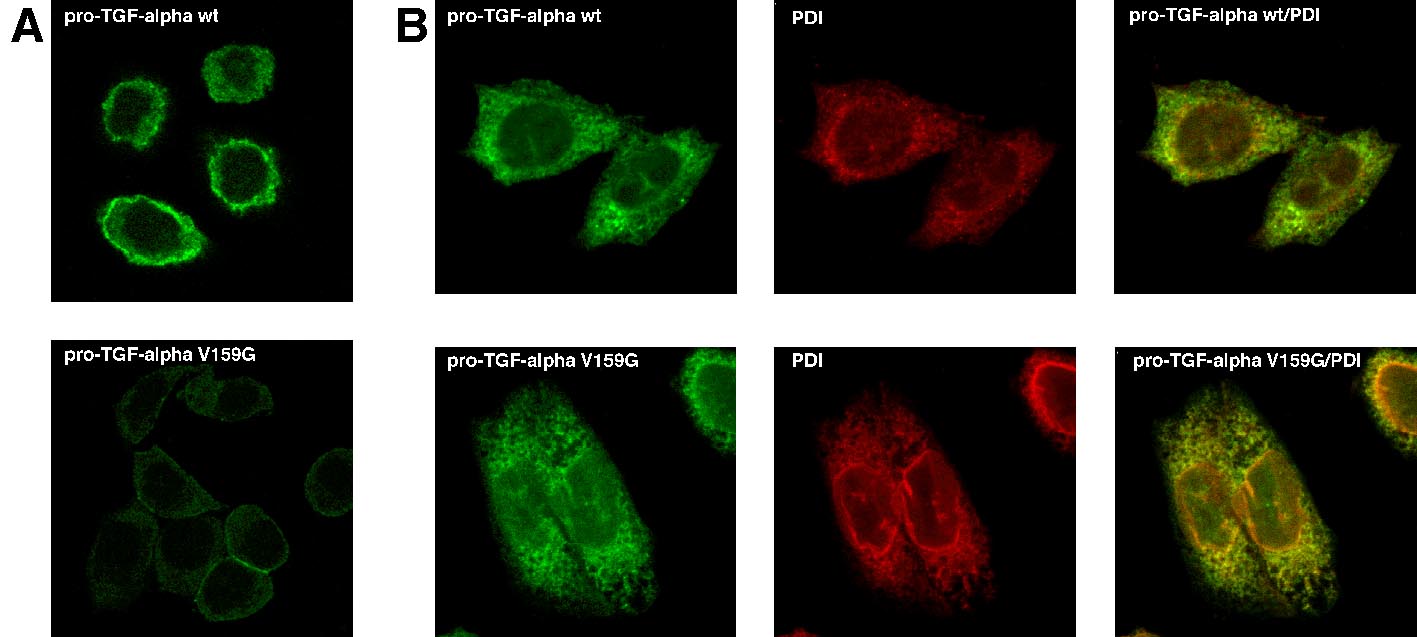
Figura 2. Localización subcelular de los mutantes del pro-TGF-alfa en el dominio carboxi-terminal. A) Células CHO (Chinese Hamster Ovary) provenientes de ovario de hamster, fueron transfectadas con pro-TGF-alfa salvaje (pro-TGF-alfa wt) o mutante (pro-TGF-alfa V159G), después fueron incubadas con anticuerpos dirigidos contra el TGF-alfa unidos a fluoresceína. La distribución del pro-TGF-alfa se detecta en forma de color verde y se ve menor marcaje de superficie en la forma mutante con respecto a la forma salvaje. B) Las mismas células se incubaron con anticuerpos dirigidos contra el TGF-alfa unidos a fluoresceína y anticuerpos dirigidos con la proteína disulfido isomerasa (PDI) unidos a rodamina. La proteína PDI es específica de Retículo Endoplasmático. La distribución de pro-TGF-alfa se ve en verde, la localización de PDI en rojo, la imagen combinada binaria de la derecha muestra la colocalización de ambas proteínas en forma de flourescencia intermedia de color amarillo, demostrando la retención de la forma mutante a nivel de RE.
Saturabilidad y Generalidad del Mecanismo de Transporte
Estudiamos a continuación la saturabilidad del mecanismo de transporte, con lo que pudimos ver que el mismo TGF-alfa era capaz de competir, desplazando la llegada a la superficie celular de nuestra proteína. Sin embargo, la proteína betaglicano, cuyo transporte a la membrana no se ve afectada por las mutaciones de la zona intracelular, no era capaz de competir o disminuir la cantidad de TGF-alfa capaz de llegar a la membrana. Esto indica que su transporte tendría lugar por un mecanismo diferente, y con el que no interferiría el TGF-alfa. Es interesante destacar que tampoco compite significativamente un mutante que no es transportado a la membrana (TGF-alfa con la valina C-terminal mutada), lo que nos indica que existe un componente molecular que discrimina entre las moléculas funcionales y las mutadas, y que solamente es saturado por las moléculas naturales. Al no poder interactuar con la molécula mutada no puede saturar el mecanismo de transporte.
Para determinar si la existencia de una valina C-terminal es una señal general de transporte a membrana comprobamos que existen más proteínas de membrana que también presentan este aminoácido en el extremo C-terminal. Un ejemplo son las metaloproteasas de membrana, proteínas integrales de membrana, que llevan a cabo funciones de ruptura proteica, contribuyendo a la activación o inactivación de enzimas extracelulares y que están ancladas en la bicapa lipídica. Realizando un abordaje similar de mutagénesis dirigida de las proteínas pudimos comprobar que cuando mutábamos la valina terminal de estas moléculas a glicina, el transporte a la superficie también quedaba alterado y se acumulaba también en el RE, al igual que ocurría con la mutación del TGF-alfa.
Nuestros resultados, por lo tanto, sugieren un sistema de transporte saturable necesario para la asignación correcta de ciertas moléculas de transmembrana a la superficie celular. Sugieren, adem< s, que para la interacción con dicho sistema es crucial la identidad del aminoácido carboxi-terminal de las moléculas clasificadas, i.e., una mutación de un único aminoácido en el extremo de una proteína impide que J sta se dirija correctamente a su destino y hace que se acumule, por lo tanto, en un orgánulo que hace una función de control de calidad. La pregunta que nos aparece ahora es: ¿cu< les son los componentes moleculares que interaccionan, bien con la proteína mutante o bien con la proteína normal, y que nos produce el efecto biológico que observamos?
Identificación de un componente de la maquinaria molecular de transporte a la superficie
Para responder a esta cuestión, hemos utilizado un abordaje que consiste en la detección de proteínas de interacción mediante el sistema de "doble híbrido" en levadura (Ausubel, 1987). Con esta técnica, la expresión de un gen que permite la supervivencia de las levaduras en un medio de cultivo deficiente en algún nutriente, queda bajo el control de un promotor o región reguladora de un gen que únicamente se activa mediante la formación de un complejo proteico entre la proteína que estamos analizando y las posibles proteínas que puedan interaccionar o formar complejos estables con ellas. De esta manera es posible aislar e identificar proteínas desconocidas o proteínas cuya función no es conocida. Realizamos este abordaje con la proteína normal TGF-alfa y lo comparamos con la proteína mutada en la cual la valina terminal había sido cambiada a glicina. Esta metodología nos permitió identificar un total de 76 proteínas que interaccionaban con la proteína TGF-alfa, es decir, que podían formar un complejo estable proteína-proteína en el interior de la célula de levadura. De estas 76 proteínas, había cinco que no eran capaces de interaccionar con la molécula TGF-alfa mutada en la valina terminal a glicina, lo que indicaba que posiblemente se tratara de moléculas implicadas en el proceso que habíamos detectado, según el cual el transporte de las moléculas mutadas se veía afectado y en lugar de ser llevadas a la membrana eran acumuladas en el RE. La secuenciación a nivel de DNA de los clones aislados y la correspondencia en secuencia de proteína, nos dieron como resultado que los cinco clones eran en realidad dos que hemos denominado TACIP-1 y TACIP-18. Ambas moléculas contienen regiones conservadas que se encuentran en otras proteínas que forman complejos multiproteicos denominados PDZ. Hemos visto que ambas moléculas se unen no solamente en células de levadura sino también en condiciones fisiológicas al TGF-alfa formando un complejo estable. Además, mutaciones que distorsionan el transporte a la superficie también impiden la unión con TACIP-18 lo que apunta hacia una participación muy importante de esta molécula en el proceso de transporte.
Paralelamente, Grootjans et al.(1997) habían descrito que la misma proteína TACIP-18 era capaz de interactuar con otra proteína funcional y estructuralmente distinta del TGF-alfa: Sindecano-2. De cara a demostrar la importancia fisiológica de la interacción TGF-alfa-TACIP-18, probamos el intercambio de la zona de interacción con TACIP-18 del TGF-alfa con la zona de interacción con TACIP-18 del Sindecano-2, mediante la construcción de moléculas quimera que contuvieran la zona extracelular del TGF-alfa y la zona intracelular del sindecano. Pudimos ver que, efectivamente y de acuerdo con lo esperado, el transporte de TGF-alfa no era alterado por el cambio, detectándose la molécula quimera en la superficie celular en cantidades comparables a la molécula no alterada, lo que demostraba que el proceso clave en el transporte de esta molécula era la interacción con TACIP-18.
Cuando analizamos la distribución intracelular de TACIP-18, vimos que, al contrario de lo esperado para una proteína con dominios conservados PDZ, esta proteína se acumulaba a nivel de RE como puede observarse en la Figura 3. Esto queda demostrado por la colocalización que detectamos en forma de fluorescencia amarilla, fruto de la emisión conjunta de fluorescencia del marcador de RE: PDI y TACIP-18. Asimismo, pudimos ver que también existía colocalización entre TGF-alfa y TACIP-18 tal y como era de esperar y que esta colocalización ocurría a nivel de RE.
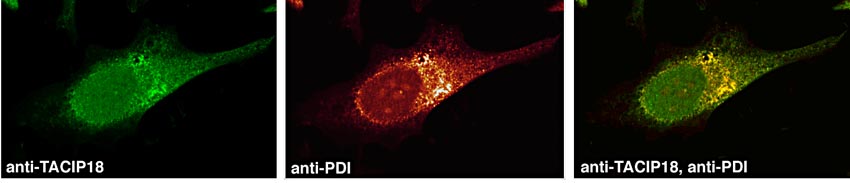
Figura 3. Localización subcelular de TACIP18. Células Hela (humanas, inmortalizadas), se incubaron con anticuerpos anti-TACIP18 unidos a fluoresceína y anticuerpos anti-PDI (proteína específica de Retículo Endoplasmático) unidos a rodamina. En verde puede verse la distribución de TACIP18 (fluoresceína) y en rojo la distribución de PDI (rodamina). La imagen binaria combinada de la derecha muestra, en forma de fluorescencia intermedia de color amarillo la colocalización correspondiente entre TACIP18 y PDI.
En resumen, el trabajo desarrollado hasta la fecha, nos hace pensar que importantes interacciones moleculares tienen lugar a nivel del RE y posiblemente del AG entre la molécula transportada y clasificada y la maquinaria molecular responsable de esta función. Empezamos a conocer algunos de los componentes moleculares que participan en este proceso, de los cuales TACIP-18 es seguramente uno de los primeros en ser identificados, y que tal vez interaccionará directa o indirectamente con los coatómeros. Probablemente existirán más y la secreción y transporte de proteínas a la membrana plasmática no sea un proceso que tiene lugar automáticamente sino que es activamente regulado por la célula. Asistiremos en el futuro próximo al esclarecimiento de una vía esencial para la fisiología de la célula.
Bibliografía
Amara, J.F. Cheng, S.H. & Smith, A.E. (1992) Intracellular protein trafficking defects in human diseases. Trends Cel. Biology, 2: 145-149.
Arribas, J., Lopez-Casilas, F. & Massagué, J. (1997). Role of the juxtamembrane domains of the transforming growth factor-alpha precursor and the beta amyloid precursor protein in regulated ectodomain shedding. J. Biol. Chem., 272(27): 17160-5.
Arribas, J., Coodly, L., Vollmer, P., Kishimoto, T.K., Rose-John, S. & Massague, J. (1996). Diverse cell surface protein ectodomains are shed by a system sensitive to metalloprotease inhibitors. J. Biol. Chem., 27(19): 11376-82.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K. (1987). Current Protocols in Cell Biology. John Wiley and Sons. New York.
Blobel, G. & Dobberstein, B. (1975). Transfer of proteins accross membranes. J. Cell. Biol., 67: 835-51 .
Bosemberg, M.W., Pandiella, A. & Massagué, J. (1992). The cytoplasmic carboxy-terminal amino acid specifies cleavage of membrane TGF-alfa into soluble growth factor. Cell, 71: 1157-1165.
Briley, G.P., Hissong, M.A., Chiu, ML., & Lee, DC. (1997). The carboxy-terminal residues of pro-TGF-alpha are required for its efficient maturation and intracellular routing. Mol. Biol. Cell, 8: 1619-31.
Fernández-Larrea, J., Merlos-Suárez, A., Ureña, J.M., Baselga, J. & Arribas, J. (1999). A role for a PDZ protein in the early secretory pathway for the targeting of pro-TGF-alfa to the cell surface. Molecular Cell, 3: 1-20.
Grootjans, J.J., Zimmermann, P., Reekmans, G., Smets, A., Degeest, G., Durr, J. & David, G.L (1997). Syntenin, a PDZ protein that binds syndecan cytoplasmic domains. Proc. Nat. Acad. Sci USA, 94: 13583-13688.
Nishimura, N., Balcj, W.E. (1997). A di-acidic signal required for selective export from the endoplasmic reticulum. Science, 277, 556-558.
Pelham, H.R. (1989). Control of protein exit from endoplasmic reticulum. Ann. Rev. Cell. Biol., 5: 1-23.
Rothman, J.E.(1994). Mechanisms of intracellular protein transport. Nature, 372: 55-63.
Sudhof, T.C. (1995). The synaptic vesicle cycle: a cascade of protein-protein interactions. Nature, 375: 645-53.
Teasdale, R.D. & Jackson, M.R. (1996). Signal mediated sorting of membrane proteins between the endoplasmic reticulum and the Golgi apparatus. Ann. Rev. Cell. Dev. Biol., 12: 27-54.
Ureña, JM., Merlos-Suárez, A., Baselga, J. & Arribas, J. (1999). The cytoplasmic carboxy-terminal amino acid determines the subcellular localization of pro-TGF-alfa and membrane type matrix metalloprotease (MT1-MMP). J. Cell. Sci., 112: 773-784.
