
QUIMICA |
Fotosíntesis y Cambios en la Composición de la Atmósfera |
Hoy en día resulta obvio que la vegetación ejerce una importante influencia sobre su medio ambiente físico. Así, la cobertura vegetal modifica el balance de radiación de la superficie terrestre al alterar el albedo, afecta al régimen de lluvias en una determinada región o cambia las características del suelo en el que crece. Dado que estos efectos modulan el crecimiento de la propia vegetación, este proceso de interacción puede ser calificado como de retroalimentación o "feed back" (Raven 1998a). Un ejemplo de dicho proceso a escala planetaria podría ser el aumento de la concentración del CO2 atmosférico producido durante las últimas décadas. Como es sabido, el CO2 es el principal gas causante del conocido como efecto invernadero, cuya consecuencia más dramática podría ser el calentamiento global de la atmósfera con el consiguiente cambio climático. Aunque la causa última de la acumulación del CO2 no es la vegetación, sí parece ser que ésta es la responsable de se esté produciendo a un ritmo menor del que podría hacerlo. Esta hipótesis se basa en que parte del "exceso" de CO2 emitido por la actividad humana, podría ser fijado por la vegetación mediante un aumento de sus tasas de fotosíntesis (en definitiva, de sus tasas de crecimiento). Dado que las tasas de fijación de CO2 varían de una especie a otra, se espera que la contribución de cada una de éstas al citado efecto amortiguador sea diferente. Por lo tanto, la hipótesis enunciada lleva a una segunda hipótesis, tal es que el mayor aumento en las tasas de crecimiento de unas especies frente a otras podría producir cambios en la estructura de los ecosistemas vegetales. Una buena forma de abordar esta pregunta es desde una perspectiva global (si bien partiendo de una escala temporal totalmente diferente), y que consiste en analizar cómo han coevolucionado atmósfera y vegetación desde la aparición de los primeros organismos fotosintéticos. La fotosíntesis es un proceso complejo en el que se implican diferentes compartimentos celulares y mecanismos enzimáticos. Además, es un proceso fuertemente dependiente de las variables físico-químicas del medio. Es debido a esta dependencia que la vegetación (y sus restos fósiles) se encuentra tan íntimamente ligada al clima (o paleoclima) de una determinada región.
Siguiendo el mismo principio, se podría esperar que los cambios producidos durante la evolución en la compleja "maquinaria fotosintética", reflejen los cambios producidos en la composición de la atmósfera (Raven 1998b).
Los registros fósiles indican que la composición de la atmósfera terrestre ha variado sustancialmente a lo largo de las diferentes eras geológicas (Berner 1990, 1993, 1994). Algunos de dichos cambios han ido ligados a la evolución de la actividad biológica sobre el planeta. En particular, los organismos fotosintéticos capaces de utilizar H2O como fuente de electrones, con el consiguiente desprendimiento de O2, son los responsables de la acumulación de éste en la atmósfera actual, dado que, según Holland (1984), el principal proceso no biológico capaz de producir O2, la fotodisociación del vapor de H2O, sólo pudo dar lugar a un aumento de la concentración de este gas de menos de 10-8 veces la actual. Las primeras células capaces de realizar fotosíntesis oxigénica fueron cianobacterias que aparecieron hace unos 3450 millones de años (Ma) (Berbee y Taylor 1993, Kooistra y Mellin 1996, Raven 1995). Antes de la aparición de estos organismos, el O2 estaba presente en la atmósfera sólo en muy pequeñas cantidades. Durante el Proterozoico (hace 2000 Ma), y coincidiendo con la aparición de los primeros organismos vegetales con estructura parecida a una alga, este gas empezó a acumularse en la atmósfera. Hacia el final de este período, se alcanzaron niveles similares al actual, coincidiendo con la aparición de los primeros ecosistemas terrestres vegetales (que datan de hace unos 1200 Ma). Posteriormente se han producido oscilaciones de la concentración de O2, alcanzándose muy altos niveles en el Carbonífero, coincidiendo con la probable aparición de las haptofitas, los ancestros de las embriofitas (cuyos primeros restos fósiles datan de 428 Ma). Paralelamente a estos eventos evolutivos, el CO2 ha sufrido también variaciones atribuibles en parte a su consumo por los organismos fotosintéticos. Así, y en contraste con la variación del contenido de O2, la concentración de CO2 en la atmósfera ha disminuido desde niveles superiores a 3 kiloPascal (kPa) hace 3500 Ma, hasta su nivel actual (unos 30 Pa).
Los cambios en la composición de la atmósfera durante la evolución de la Tierra tuvieron importantes consecuencias sobre las condiciones de habitabilidad en ella. Siguiendo con el O2, su acumulación supuso el paso de una atmófera neutra a ligeramente reductora, lo que afectó el ciclo biogeoquímico de otros nutrientes esenciales. Así, se produjo la oxidación del Fe(II) a Fe(III) con la consiguiente precipitación de este como óxido (Couturect et al. 1994). La presencia de fases sólidas de Fe(III), capaces de enlazar fósforo en forma de ortofosfato, produjo una limitación en la disponibilidad de este último elemento. También se favoreció la aparición de nitrógeno combinado así como la inhibición de la fijación biológica del N2. La acumulación de O2 también condujo a la formación de la capa de ozono estratosférica, con importantes consecuencias en la protección frente a la radiación ultravioleta (Rozema et al. 1997).
Sin duda, la consecuencia más directa de los cambios en la concentración de O2 y CO2 sobre los organismos fotosintéticos, pudieron ser variaciones en las tasas de carboxilación de la Rubisco. Como es sabido, la Rubisco es la principal enzima encargada de la fijación fotosintética del CO2, lo que determina finalmente las tasas de crecimiento vegetal. El O2 compite con el CO2 por el sitio activo del enzima, de manera que las tasas de carboxilación dependen de la concentración de CO2 relativa a la de O2 así como de la selectividad del enzima para el CO2 frente al O2. Altos valores del índice CO2/O2 suprimen la actividad oxigenasa de la Rubisco. Por el contrario, cuanto menor sea dicho índice, más limitada podría estar la fotosíntesis por el CO2. Así, y a modo de ejemplo, la concentración de CO2 atmosférico actual no es suficiente para suprimir la actividad oxigenasa de la Rubisco y saturar las tasas de carboxilación. Los principales grupos de plantas presentes en la actualidad surgieron bajo ambientes con muy diferentes valores del índice CO2/O2. Las primeras cianobacterias con fotosíntesis oxigénica debieron realizar la fotosíntesis bajo una atmósfera rica en CO2 y carente prácticamente de O2. Durante la aparición de las primeras células fotosintéticas eucariotes, el valor del índice CO2/O2 pudo ser aproximadamente unas 10 veces mayor al actual. El desarrollo posterior de la vegetación (en especial la aparición de la vegetación embriofítica terrestre) se produjo bajo valores del índice similares al actual, aunque hubo importantes oscilaciones (principalmente en el Carbonífero).
Hoy en día está bien establecido que los grupos vegetales tienen afinidades diferentes por el CO2 debido a (1) las propias características cinéticas de la Rubisco y (2) la presencia de mecanismos de concentración de carbono (MCC) cuya función es aumentar la concentración de CO2 en el entorno del enzima, favoreciendo así la actividad carboxilasa y suprimiendo la actividad oxigenasa. La Rubisco de las cianobacterias tiene una menor selectividad para el CO2 frente al O2 que la de las algas verdes y plantas terrestres. Los valores mayores de selectividad para el CO2 se encuentran, sin embargo, entre algas rojas y pardas. La Rubisco de las algas verdes y embriofitas fue adquirida directamente de una cianobacteria mientras que la de las algas rojas y pardas fue derivada de otro grupo primitivo de bacterias (Bhattacharye y Medlin 1995). Así pues, los diferentes valores de selectividad de la Rubisco de estos organismo podrían resultar de su diferente origen filogenético así como de su desarrollo evolutivo posterior. Además, ambas moléculas primitivas, surgidas bajo ambientes ricos en CO2, debieron evolucionar en paralelo hacia mayores valores de selectividad para el CO2 cuando el nivel de este declinó en la atmósfera. Por lo tanto,
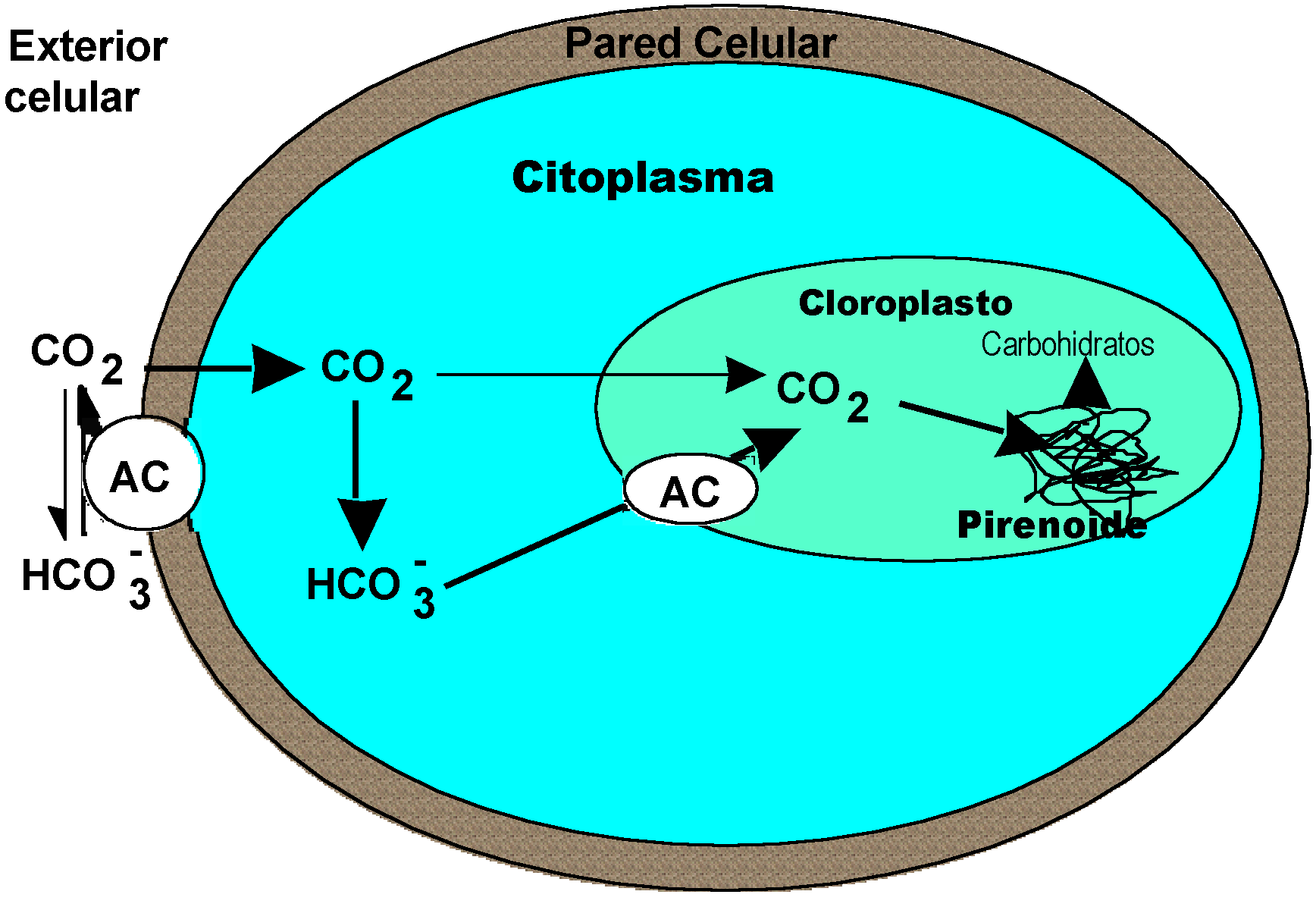
Figura 1. Esquema de una célula de un alga eucariote marina mostrando la ubicación probable y la función de la AC. En el agua de mar, el pH es aproximadamente 8.1 y la concentración de carbono inorgánico total es 2.2 mM. Debido al pH alcalino del agua de mar aproximadamente el 95% del carbono inorgánico se encuentra como HCO3-que no puede difundir libremente hacia el interior de la celula. La presencia de AC en el exterior de la célula permite que el HCO3- se transforme rápidamente en CO2 que puede entrar en la célula por difusión. En el interior el CO2se almacena como HCO3-. La AC interna facilita el transporte del HCO3- almacenado hacia el interior del cloroplasto. Como resultado, se mantienen una alta concentración de CO2 alrededor de la Rubisco, saturándose sus tasas de carboxilación y reduciéndose su actividad oxigenasa.
La afinidad de la Rubisco por el CO2 en algunos grupos de plantas podría reflejar las características de la atmósfera bajo la que surgieron. Sin embargo, como se ha señalado, además de la Rubisco, muchas cianobacterias, algas y plantas terrestres presentan mecanismos de concentración de CO2 que están estrechamente ligados a la actividad de esta enzima. Existen dos tipos de mecanismos: (a) basados en el transporte de carbono inorgánico (CO2 o HCO3-) a través de las membranas (presentes en plantas acuáticas, principalmente en cianobacterias y algas) y (2) basados en ciclos de carboxilación/descarboxilación que preceden la acción de la Rubisco (presentes en plantas denominadas C4 y CAM según el tipo de ruta fotosintética que posean). El origen polifilético de estos mecanismos es generalmente aceptado (Apel 1994), si bien el momento evolutivo en que pudieron aparecer sigue siendo materia de controversia. No obstante, las rutas C4 y CAM no deben ser más antiguas, ya que surgieron hace 200 Ma con niveles de CO2 y O2 similares al actual (esto es, concentraciones de CO2 no saturantes para la fotosíntesis). Sin embargo, los MCC basados en el transporte del carbono inorgánico a través de membranas, debieron surgir mucho antes de que el nivel de CO2 o la razón CO2/O2 hayan podido favorecer su selección, ya que las cianobacterias datan de 3500 Ma y los macrófitos acuáticos de hace unos 600 Ma. Un componente esencial de los MCC basados en el transporte a través de membrana es la enzima denominada anhidrasa carbónica (AC). Esta enzima se encuentra ampliamente distribuida en todos los grupos vegetales, desde las cianobacterias hasta las plantas vasculares terrestres. Su función es la de acelerar la transformación reversible del CO2 a HCO3-. La presencia de AC en la superficie de las células de plantas acuáticas permite el uso de HCO3- como fuente de carbono inorgánico para la fotosíntesis al acelerar su transformación en CO2, que puede difundir fácilmente hacia el interior de la célula (Figura 1). Dado que la concentración de HCO3- en el agua de mar es unas 200 veces mayor que la de CO2 (aproximadamente 12 microM), la presencia de AC externa en plantas acuáticas supone el acceso a una fuente mayor de carbono inorgánico que la que tendría si sólo pudiera usar este CO2. Como consecuencia, las tasas de fotosíntesis de muchas de estas plantas están saturadas a la concentración de carbono inorgánico presente en su medio. En cianobacterias y algas con CCM la presencia de AC es, además, necesaria en el citoplasma y estroma del cloroplasto, puesto que el carbono inorgánico se almacena en el interior como HCO3-, que ha de ser transformado posteriormente en CO2 para que pueda ser fijado por la Rubisco (Suzuki et al. 1994). Diferentes estudios experimentales llevan a la conclusión de que la AC interna no es precisa cuando la concentración de CO2 externa es alta (Mercado et al. 1997a,b), esto es, la AC no tiene función fotosintética aparente en una atmósfera con elevada concentración de CO2 (como debió ser la atmósfera bajo la que surgieron las cianobacterias y algunos de los principales grupos de algas). Otro componente de los MCC basados en el transporte es el pirenoide, una estructura proteinica que se encuentra en el interior del cloroplasto y se supone ligada estrechamente a la actividad de la Rubisco. Los registros fósiles indican que estructuras celulares tipo pironoide aparecieron ya en eucariotas con 1000 Ma de antigüedad, esto es, en una atmósfera con una concentración de CO2 unas 40 veces mayor que la actual. Por lo tanto, los datos disponibles indican que algunos componentes de los mecanismos de concentración de carbono basados en el transporte de carbono inorgánico no surgieron bajo una atmósfera global con elevados "déficit" de CO2.
En resumen, la afinidad para el CO2 en algunos grupos de plantas podría reflejar las características de la atmósfera global bajo la que surgieron. Sin embargo, estas características ambientales globales no justifican la aparición y su selección por evolución de algunos mecanismos fotosintéticos, como los MCC basados en el transporte de carbono inorgánico a través de membranas. Aunque las razones para explicar esta "falta de correlación" son obviamente materia de especulación, el origen temprano de algunos MCC puede deberse a que el CO2 pudo limitar la fotosíntesis mucho antes de que su concentración en la atmósfera descendiera. Así por ejemplo, los microorganismos fotosintéticos bentónicos que crecen en mantos microbianos bajo el agua o en tierra (Raven 1993) e incluso algunos macrófitos acuáticos (Mercado et al. 1998) podrían llegar a estar limitados por el CO2 debido a la "lenta" velocidad de difusión de este gas unida a una disminución de su concentración al ser consumido por el propio organismo. Nótese que al aludir a distancias de difusión y concentraciones locales se ha de pasar de considerar la interacción planta-medio ambiente desde una escala global a una micro- o mesoescala, poco dependiente en principio de las características medias de la atmósfera. Este hecho lleva a reorientar la cuestión que dio partida a esta disertación, a saber, en qué medida podrían responder los ecosistemas vegetales terrestres al aumento relativamente vertiginoso de la concentración del CO2 atmosférico: debido a la importancia de las condiciones ambientales "locales" es muy probable que el aumento del CO2 atmosférico que se está produciendo en la actualidad tenga poco efecto sobre algunos ecosistemas vegetales, principalmente sobre los ecosistemas vegetales acuáticos. No obstante, dada la escasez de datos y de estudios adecuadamente orientados, es preciso aún mucho trabajo experimental y de campo para poder emitir alguna predicción suficientemente fundada.
Referencias
Apel P. 1994. Evolution of the C4 photosynthetic pathway: a physiologists point of view. Photosynthetica 30: 495-502.
Bhattacharya D. & Merlin L. 1995. The phylogeny of plastids: a review based on comparisons of small-subunit ribosomal RNA coding regions. J. Phycol 31: 489-498.
Berbee M. L. & Taylor J. W. 1993. Dating the evolutionary radiations of the true fungi. Can. J. Bot. 71: 1114-1127.
Berner R. A. 1990. Atmospheric carbon dioxide levels over Phanerozoic time. Science 249: 1382-1386.
Berner R. A. 1993. Paleozoic atmospheric CO2: importance of solar radiation and plant evolution. Science 261: 68-70.
Berner R. A. 1994. Geocarb II: a revised model for atmospheric oxygen over Phanerozoic time. Am. J. Sci. 289: 331-361.
Couture M., Chamberland H., St-Pierre B. & Guertin M. 1994. Nuclear genes encoding chloroplasts hemoglobins in the unicellular green alga Chlamydomonas eugametos. Mol. Gen. Genet. 243: 185-197.
Holland H. D. 1984. The Chemical Evolution of the Atmosphere and Oceans. Princenton University Press, Princenton, N. J.
Kooistra W.H.C.F. & Mellin L. 1996. The origin of the pigmented heterokonts and diatoms. J. Phycol. 32: 25s
Mercado J. M., Figueroa F. L. & Niell F. X. 1997a. Regulation of the mechanism for HCO3- using by the inorganic carbon level in Porphyra leucosticta. Planta 201: 319-325.
Mercado J. M., Niell F. X., Figueroa F. L. & Axelsson L. 1997b. A new method for estimating external carbonic anhydrase activity in macroalgae. J Phycol 33: 999-1006.
Mercado J. M., Gordillo F. J. L., Figueroa F. L. & Niell F. X. 1998. External carbonic anhydrase in tidal macroalgae. J Exp Mar Biol Ecol 221: 209-220.
Raven J. A. 1993. The evolution of vascular land plants in relation to quantitative function of dead water-conducting cells and stomata. Biol. Revs. 68: 337-363
Raven J. A. 1995. The early evolution of land plants: aquatic ancestors and atmospheric interactions. Bot. J. Scotland 47: 151-175.
Raven J. A. 1998a. Extrapolating feedback processes from the present to the past. Phil. Trans. R. Soc. Lond. B. 353, 19-28.
Raven J.A. 1998b. Phylogeny, paleoatmosphera and the evolution of phototrophy. Stable isotopes (ed. H. Griffiths). BIOS Scientific Publishers Ltd., Oxford.
Rozema J., van de Staiij J., Björn L. O. & Cadwell M. 1997. UV-B as an enviromental factor in plant life: stress and regulation. Trends Ecol. Evoln. 12, 22-28.
Suzuki E., Shirawa Y. & Miyachi S. 1994. The cellular and molecular aspects of carbonic anydrase in photosynthetic organisms. Progress in Pycological Research. Vol 10 (F. E. Round, D. J. Chapman, eds).
