
BIOLOGIA  |
Dimorfismo Sexual en el Cerebro |
|
|
© Juan
Carlos Jorge-Rivera, 1998
|
La naturaleza provee los mecanismos anatómicos y endocrinos
que aseguran la reproducción de las especies. Durante el periodo
prenatal las hormonas sexuales organizan la estructura de nuestros cuerpos
y nos asigna un sexo, macho o hembra. Durante la pubertad las
hormonas sexuales median la aparición de características
secundarias asociadas al sexo y nos proveen con la capacidad de reproducción.
Sin embargo, las hormonas sexuales también proveen sexo al órgano
más complejo de nuestros cuerpos: el cerebro. ¿Cuáles
son algunas de estas diferencias? ¿Qué impacto tienen
éstas diferencias en la expresión de conductas sexuales?
¿En qué maneras la organización dimórfica de
estos centros cerebrales media las expresiones polimórficas de nuestra
sexualidad? Estas son algunas de las preguntas que me propongo contestar
a través de este artículo.
Dimorfismo sexual se refiere principalmente a diferencias en
el tamaño del núcleo cerebral o circuito bajo estudio. Estas
diferencias en tamaño sin embargo, se pueden deber a diferencias
en el volumen o área que el núcleo ocupe, así como
a diferencias en el número de células (o densidad) del núcleo.
Estas diferencias neuroanatómicas se asocian a sexo (macho o hembra),
preferencia sexual (homosexual o heterosexual), o transexualismo (identidad
de género correspondiente al sexo opuesto). La mayoría de
las regiones sexualmente dimórficas del cerebro pertenecen al sistema
límbico (Dulce Madeira y Lieberman, 1995).Este sistema también
es conocido como el cerebro primitivo ya que emerge desde bien
temprano en la escala evolutiva.Se le considera un sistema ya que los
circuitos o núcleos que lo componen están altamente interconectados
entre sí de manera que la actividad eléctrica y química
de un núcleo puede influenciar la actividad de otro núcleo.
No resulta sorprendente que la mayoría de los circuitos dimórficos
del cerebro sean parte o estén íntimamente relacionados con
el hipotálamo (Figura 1).
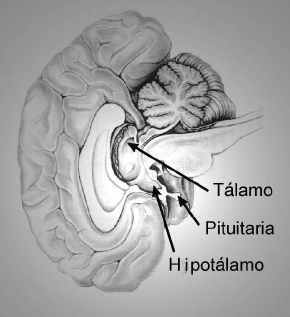 |
| Figura 1. Localización del hipotálamo en el cerebro humano.La base del cerebro (tope de la espina dorsal) se encuentra hacia la derecha y la parte posterior hacia arriba en la figura. (Modificado de Principles of Neural Science (1991), Kandel, Scwhartz, Jessel;(Editores). Conneticut: Appleton & Lange). |
El hipotálamo, entre otras funciones, regula las hormonas
y representa una interfase anatómica y funcional entre el sistema
nervioso y el sistema endocrino. De hecho, la mayor concentración
de receptores de hormonas sexuales (andrógenos, estrógenos,
y progesteronas) del cerebro se encuentran en el hipotálamo ( revisado
en Pfaff y Schwartz-Giblin, 1988). El hipotálamo tradicionalmente
se conoce como el centro ejecutivo de funciones homeostáticas del
cuerpo. En adición al control de hormonas (sexuales y no sexuales),
el hipotálamo regula patrones de hambre, sed, sueño, temperatura
del cuerpo, y conducta sexual. Como es de imaginar, la estructura
del hipotálamo es compleja tanto en términos neuroanatómicos
como neuroquímicos. Este se puede subdividir en 4 regiones
y en más de 25 núcleos de acuerdo a criterios anatómicos
y funcionales. Los núcleos dimórficos que se describen
en este artículo incluyen: el área preóptica, el núcleo
ventromedial, el núcleo supraquiasmático ( todos éstos
son subdivisiones del hipotálamo), la amígdala, y la stria
terminalis.
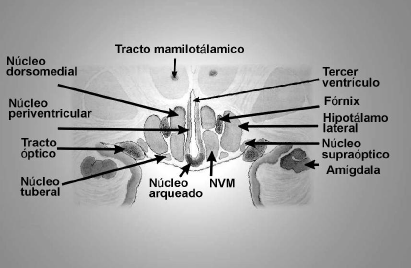 |
| Figura 2. Localización del NVM y la amígdala en un plano coronal del cerebro humano al nivel del tercer ventrículo. El NSQ se encuentra en un plano más anterior, a la base del tercer ventrículo mientras que el APO también se encuentra aun plano más anterior en el área correspondiente al hipotálamo lateral y NVM. El BNST se encuentra en un plano más posterior entre la amígdala y el tracto óptico. (Figura modificada de Principles of Neural Science, ibid). |
1. El Area Preóptica (APO)
El APO fue el primero en describirse como sexualmente dimórfico y continúa siendo uno de los ejemplos más claros de dimorfismo tanto en animales de laboratorio como en humanos (Gorski y colaboradores, 1978; Swaab y Fliers, 1985). Originalmente se le designó como el núcleo sexualmente dimórfico del área preóptica (o SDN of the POA por sus sílabas en inglés). El nombre de la región varía dependiendo de la especie, y en algunos casos se debate si la correspondencia en términos de posición anatómica es suficiente para aceptar regiones de diferentes especies como equivalentes. En ratas, el área se le conoce como la porción medial del área preóptica (mPOA por sus sílabas en inglés) y en humanos como el núcleo intersticial del hipotálamo anterior-1 (NIHA-1 o INAH-1 por sus sílabas en inglés) (Allen y colaboradores, 1989). En adición al NIHA-1 se ha descrito NIHA-2 al 4 como regiones dimórficas. Sin embargo, debido a la proximidad anatómica de estas islas de neuronas, se debate si en realidad NIHA 1-4 representan grupos neuronales anatómica y funcionalmente independientes unas de las otras. LeVay (1991) confirmó dimorfismo sexual en el NIHA-3 pero no en NIHA 1, 2, y 4. Este mismo estudio también reporta dimorfismo en el NIHA-3 con relación a preferencia sexual. El NIHA-3 es mayor en hombres heterosexuales comparado con hombres homosexuales y mujeres (LeVay, 1991). Aún se debate la validez de estos resultados ya que algunas muestras de tejido nervioso provinieron de pacientes con SIDA y el número de casos de SIDA no estaban igualmente representados en los diferentes grupos experimentales. En el caso del NIHA-1, sin embargo, se ha determinado que en los hombres existen dos veces más células que en las mujeres (Swaab y Hoffman, 1988). Incluso se ha determinado que tanto niños como niñas poseen el mismo número de células al nacer y que este número aumenta exponencialmente hasta los 4-5 años de edad. La cantidad de células permanece constante hasta la pubertad pero en el caso de las niñas un gran porciento de las células muere (Swaab y colaboradores, 1988). Por lo tanto, el dimorfismo sexual del área preóptica en humanos no es aparente hasta la pubertad.
¿Qué implicaciones tiene que el área preóptica de los machos sea mayor que el de las hembras? Ya que teóricamente no existe una correlación lineal entre el número de neuronas de un circuito y su capacidad computacional, se desconoce si diferencias en tamaño reflejan diferencias cualitativas y/o cuantitativas en términos de función neural. Modelos animales sin embargo, han sido útiles para determinar qué conductas controlan el área preóptica. En este tipo de experimento la estrategia consiste en destruir el circuito neuronal de interés, ya bien sea quirúrgicamente o por medio de compuestos tóxicos, y luego observar determinada conducta del animal en comparación con animales intactos. Es claro que si se destruye el área preóptica, las ratas machos no expresan conducta sexual en presencia de una hembra receptiva (Hansen y colaboradores, 1982; Heiman y colaboradores, 1966/1967; Szechtman y colaboradores, 1978). En las ratas hembras, el área preóptica es considerado como un circuito inhibidor de receptividad sexual (Pfaff y Schwartz-Giblin, 1988). También se ha determinado el rol del APO en conductas sexuales en primates. En macacos machos, el APO está asociado con iniciación del acto sexual, erección del pene, y el periodo refractario después del acto sexual. En las hembras el APO está asociado con iniciación del acto sexual (Oomura y colaboradores, 1989).
2. Núcleo Ventromedial del Hipotálamo (NVM)
El NVM (o VMN por sus sílabas en inglés) es facilitador de conducta sexual en las hembras (Davis y colaboradores, 1979; Pfaff y Sakuma, 1979a,b). En ratas, las hembras expresan receptividad sexual por medio de una conducta refleja que se conoce como lordosis. Durante lordosis la hembra flexiona su espina dorsal en forma de arco, levanta su cabeza en un ángulo de 90 grados, levanta sus patas traseras y mueve su cola hacia un lado al ser montada por un macho para darle acceso a la cavidad vaginal. Esta conducta de receptividad sexual se expresa durante niveles altos de hormonas sexuales (estrógeno y progesterona), de lo contrario la hembra rechaza los acercamientos sexuales del macho. El caso del dimorfismo sexual en el NVM es interesante porque no se debe a diferencias en tamaño o densidad del núcleo sino a la complejidad estructural de las neuronas. Las neuronas se comunican entre sí por medio de señales químicas (neurotransmisores) a través de contactos especializados conocidos como sinapsis. En el NVM, la densidad sináptica es mayor en machos que en hembras (Miller y Aoki, 1991). Aún más interesante, el número de contactos sinápticos en la rata hembra varía a través del ciclo de estrus (el equivalente al ciclo menstrual en la mujer) (Frankfurt y colaboradores, 1990; Cohen y Pfaff, 1992). Por lo tanto el dimorfismo del NVM está claramente controlado por las hormonas sexuales, y en el caso de las hembras es regulado a lo largo de los ciclos hormonales. En el caso de primates hembras, la actividad neurofisiológica del NVM está sincronizada con el acto sexual (Oomura y colaboradores, 1989).
3. Núcleo Supraquiasmático (NSQ)
El NSQ (o SCN por sus sílabas en inglés) regula
los ritmos cíclicos o ritmos circadianos del cuerpo. EL NSQ
genera y coordina ritmos hormonales, fisiológicos, y conductuales
con una periodicidad de cerca de 24 horas. Se ha encontrado que el
NSQ difiere en forma entre hombres y mujeres; esférico en hombres
y oblongado en mujeres (Swaab y colaboradores, 1985).
El impacto funcional de este tipo de dimorfismo no se ha determinado.
Se ha sugerido que la forma de un circuito neuronal pudiese depender de
el tipo de conexiones (aferentes y eferentes) que éste establezca
con otros circuitos. De todos modos, existe una diferencia clara
en términos del volumen y el número de células del
NSQ entre hombres heterosexuales y hombres homosexuales. Swaab
y Hoffman (1990) encontraron que tanto el volumen como el número
de neuronas es el doble en el NSQ de homosexuales en comparación
con heterosexuales. La neuroquímica de este núcleo
también difiere entre homosexuales y heterosexuales; el NSQ de homosexuales
contiene el doble de neuronas inmunoreactivas a vasopresina en comparación
con heterosexuales (Swaab y Hoffman, 1990). En roedores de
la pradera, se ha demostrado que la vasopresina media conductas asociadas
a la reproducción: agresividad, territorialidad, monogamia, y conducta
paternal. Curiosamente, la vasopresina aumenta todas estas conductas
exclusivamente en machos (Young y colaboradores, 1998).
En el caso de animales de laboratorio se ha establecido que el NSQ
también está asociado con conductas sexuales.
Sördersten y colaboradores (1981) determinaron que la amplitud
de los ritmos circadianos en conducta sexual es mayor en ratas tratadas
con anti-estrógenos durante el periodo neonatal. De
igual modo, si se administran anti-estrógenos durante el periodo
prenatal y postnatal, se encuentra un aumento en un 59% de neuronas que
contienen vasopresina en el NSQ, así como conducta bisexual en
ratas. Si el tratamiento con anti-estrógenos se limita al
periodo prenatal no se observan estos cambios (Swaab y colaboradores,
1995). En términos de conducta sexual, Krujiver y colaboradores
(1993) lesionaron el NSQ en ratas machos para determinar si expresaban
conducta homosexual o bisexual pero no se encontró tal efecto.
Curiosamente, se ha observado que ratas tratadas con anti-estrógenos
expresan conducta bisexual temprano en el ciclo nocturno y conducta heterosexual
a finales del ciclo nocturno, lo cual continúa apuntando al NSQ
como importante en expresión de conducta sexual, al menos en ratas
(Bakker y colaboradores, 1993). En estos momentos no se ha
encontrado evidencia que demuestre el rol del NSQ en el establecimiento
de preferencia sexual, a pesar de que el dimorfismo es claro entre homosexuales
y heterosexuales. Al igual que en el área preóptica,
el dimorfismo del NSQ se observa entre los 4 y 12 años de edad (Swaab
y colaboradores, 1987). Este resultado implica que el dimorfismo
de este núcleo esta bajo control de las hormonas sexuales y que
factores postnatales juegan un rol en el establecimiento del dimorfismo
del NSQ como es en el caso del área preóptica.
4. Amígdala
La amígdala medial es 85% mayor en volumen en los hombres
que en las mujeres (Hines y colaboradores, 1992). La amígdala
es considerada como el centro de las emociones, y regulador de reacciones
de agresividad, miedo, y ansiedad. Recientemente se ha demostrado
que en humanos la amígdala esta asociada con aprendizaje emocional
y (Morris y colaboradores, 1998) y conducta social (Adolphs y
colaboradores, 1998). La amígdala también juega
un rol importante en conducta sexual. En el caso del Síndrome Klüver-Bucy
tanto humanos como monos expresan hipersexualidad, lo cual incluye interés
sexual por objetos inanimados. Este síndrome está asociado
con pérdida de función de la amígdala. La amígdala
es también un centro de integración de información
olfativa, lo cual juega un rol bien importante en la expresión de
conducta sexual.
La amígdala, al igual que el hipotálamo, se subdividide
en varios subnúcleos por criterios anatómicos y funcionales.
La porción anterior de la amígdala corticomedial aumenta
receptividad en ratas hembras mientras que la porción posterior
del núcleo lateral reduce receptividad (Masco y Carrer;
1980). En contraste con el NVM, se cree que la amígdala
no media directamente respuestas lordóticas en las hembras sino
que contribuye a la motivación o excitación sexual. En el
caso de las ratas machos esta posibilidad es más clara ya que se
ha demostrado que la amígdala medial contribuye a excitación
sexual (evidenciado por erección) sin contacto sexual
(Kondo y colaboradores, 1997).
5. Stria Terminalis (ST)
La Stria Terminalis (ST) es la conección eferente principal
del la amígdala (Weller y Smith, 1982). La ST es un
tracto y se puede dividir en varias regiones. La porción base
de la ST (bed nucleus of the stria terminalis, BNST) es 97% más
grande en volumen en hombres que en mujeres (Hines y colaboradores,
1992). En ratas, lesión del BNST altera la secuencia de
copulación en machos, es decir, los machos aumentan la cantidad
de penetraciones antes de lograr eyaculación y en casos severos,
no logran eyaculación a pesar de que otros aspectos del repertorio
sexual aparecen inalterados (Emery y Sachs,1976; Claro y colaboradores,
1995). En un estudio reciente se demostró que animales
con lesiones en el BNST no logran excitación sexual (Liu y colaboradores,
1997).
En humanos, se desconoce qué rol juega el BNST en conductas
sexuales. Sin embargo, se ha encontrado dimorfismo en la subdivisión
central de éste núcleo. Zhou y colaboradores (1995)
encontraron el BNST típico de mujeres en transexuales que
cambiaron su sexo de hombre a mujer. Es impresionante que este dimorfismo
con relación a identidad de género no fue influenciado por
tratamiento hormonal durante la adultez, y tampoco correlaciona con la
preferencia sexual de los sujetos. Este es el primer estudio que
reporta una estructura dimórfica del sexo femenino en individuos
genéticamente del sexo masculino.
Proyecciones experimentales: Ciencia al Día
Como hemos visto, en algunos casos ha sido posible establecer
en modelos animales qué conductas son controladas por circuitos
dimórficos. Cabe recordar que estos experimentos han sido
realizados en su mayoría en roedores y en pocos casos, en primates
no-humanos. Por necesidades experimentales, las conductas observadas
son estereotipadas; en su mayoría conductas reflejas y asociadas
a reproducción. De todos modos, tenemos información
de los dos polos del espectro: anatomía de circuitos y conductas
asociadas a estos circuitos. Experimentalmente es difícil
correlacionar la actividad neural con una conducta. Lograr dicha
correlación sería equivalente a viajar a la luna con una
sofisticada grabadora de sonido y grabar aleatoriamente los sonidos de
un individuo (neurona) entre la población digamos de una isla (circuito
dimórfico), asumiendo que el discurso del individuo (actividad neuronal),
refleje la conducta de los habitantes de la isla (conducta sexual).
Como mencionara anteriormente sin embargo, todos los circuitos dimórficos
aquí descritos están intimamente relacionados en términos
neuroanatómicos (por conecciones entre sí) y neuroquímicamente
(por compartir neurotransmisores, y ambientes hormonales muy similares
entre sí). Estas similitudes prestan una oportunidad experimental
única para caracterizar las propiedades fisiológicas que
comparten las neuronas de estos centros cerebrales.
El sistema de neurotransmisión más abundante en estos
circuitos dimórficos es el de ácido gama-amino-butírico
(GABA por sus sílabas en inglés).Transmisión GABAérgica
es predominante en todos los circuitos discutidos en este artículo:
el APO (Jarry y colaboradores, 1991; Ondo y colaboradores, 1982),
el NVM (Makara y colaboradores, 1975; OConnor y colaboradores, 1988;
Ogawa y colaboradores, 1991), el NSQ (Decavel y van del Pol, 1990;
Wagner y colaboradores, 1997), el BNST (Wisden y colaboradores,
1992), y la amígdala (Nitecka y Ben-Ari, 1987; Nitecka y
Frotscher, 1989). La función principal de GABA es inhibir
la actividad eléctrica del cerebro. Por ejemplo, algunos casos
de epilepsia severa, (donde el cerebro sufre de hiperexcitabilidad) se
asocia con defectos en el sistema de transmisión GABAérgico.
Ya que el sistema de GABA provee el mecanismo principal para controlar
la excitabilidad neuronal se hipotetiza que éste juega un papel
crítico en procesos computacionales del cerebro. En términos
de conducta sexual se ha demostrado que tanto los niveles de GABA en el
cerebro (Herbison y Fénelon, 1995; McCarthy, 1995) así
como el número de receptores de GABA, son susceptibles a hormonas
sexuales (Jüptner y Hiemke1990; Pérez y colaboradores, 1986;
Schumacher y colaboradores, 1989). En términos conductuales
se ha demostrado que la transmisión de GABA en el NVM del hipotálamo
es necesaria para la expresión de lordosis en hembras (McCarthy
y colaboradores, 1990, 1991, 1995). Recientemente se ha
demostrado que los receptores de GABA tienen función dimórfica
en el NVM; las corrientes de GABA son más rápidas en el NVM
de hembras que en machos (Smith y colaboradores, 1996). La
composición molecular del receptor determina sus propiedades
biofísicas y su susceptibilidad a agentes farmacológicos.
Algunos sedantes, tienen efecto fisiológico a través de la
modulación de los receptores de GABA. Valium por ejemplo,
hace los receptores más efectivos (i.e., Valium aumenta la inhibición
del sistema nervioso causada por GABA). Trabajo reciente en el laboratorio
de Henderson (Departamento de Fisiología, Escuela de Medicina de
Dartmouth) demuestra que existe dimorfismo sexual en cuanto a la susceptibilidad
de los receptores de GABA a calmantes que pertenecen a la familia de benzodiazepinas.
Especificamente, la droga Zolpidem reduce la eficacia de GABA en el APO
de ratas machos pero no tiene efectos fisiológicos en el APO de
ratas hembras (Nett y colaboradores, 1998). En humanos,
se ha demostrado que Valium tiene efectos reducidos durante la fase lútea
del ciclo menstrual en mujeres que padecen del Síndrome Premenstrual
(Sundström y colaboradores, 1997). De hecho, una
estrategia útil que permite caracterizar las propiedades funcionales
de las neuronas en circuitos dimórficos es determinar la susceptibilidad
de estas neuronas a diferente tipos de fármacos. Mis propias
investigaciones revelan que aún si se comparan áreas dimórficas
del mismo sexo (el NVM y el APO), las neuronas de cada circuito responden
diametralmente opuestas a esteroides sintéticos (Jorge-Rivera
y Henderson, 1998). Estos resultados no sólo apuntan a
la necesidad de considerar circuitos neurales dimórficos en el diseño
de drogas para tratar enfermedades del sistema nervioso, sino que también
demuestran por primera vez, dimorfismo funcional en sistemas de neurotransmisión
en circuitos estructuralmente dimórficos. Un acercamiento
experimental que pudiese resultar útil sería estudiar las
propiedades del sistema GABAérgico a lo largo de la cadena de estructuras
dimórficas del sistema límbico para determinar qué
contribución este sistema aporta en términos de integración
de información neural. La hipótesis principal
en este marco experimental es que conductas sexuales complejas son producto
de la interacción entre circuitos dimórficos en vez de ser
controladas por circuitos individuales.
Implicaciones:
¿Qué implicaciones tiene el que existan diferencias estructurales en el cerebro de acuerdo a género, preferencia sexual, e identidad de género? ¿Acaso son estas diferencias producto o causantes de diversas conductas sexuales en humanos? En todos los casos estudiados hasta hoy se ha determinado que el dimorfismo sexual del cerebro se hace evidente entre los 4 y los 12 años (Swaab y colaboradores, 1987, 1992). Después de todo, Freud tenía razón al establecer que la sexualidad humana no es exclusivamente programada por la biología de nuestros cuerpos sino que también es mediatizada por el ambiente (de acuerdo al psicoanálisis, por la relación del niño o niña con sus progenitores). Es posible que una multiplicidad de factores de carácter genéticos, hormonales, químicos, psicológicos, y ambientales construyan el sustrato biológico que media y permite la expresión polimórfica de nuestra sexualidad. El reto consiste en determinar, desde el modelo científico, cuál es la naturaleza de algunas de estas influencias en el establecimiento del dimorfismo sexual del cerebro.
Agradecimientos.
El autor agradece a la National Science Foundation (DBI-9707826) por proveer fondos de investigación y al Dr. Jorge Golowasch por su invaluable contribución editorial en este artículo.
Referencias
Libros.
A Separate Creation: The Search for the Biological Origins of Sexual Orientation (1996). Chandler Burr. New York: Hyperion, 354 páginas.
Gay Science: The Ethics of Sexual Orientation Research. (1997). Timothy F. Murphy. New York: Columbia University Press, 268 páginas.
Neurobiological Effects of Sex Steroid Hormones (1995). Paul E. Micevych & Ronald P. Hammer, Jr., editores. New York: Cambridge University Press, 444 páginas
The Prehistory of Sex: Four Million Years of Human Sexual Culture. (1996). Timothy Taylor. New York: Bantman Books, 353 páginas.
The Science of Desire: The Search for the Gay Gene and the Biology of Behavior. (1995). Dean Hamer & Peter Copeland. New York: Touchtone, 272 páginas.
Science in the Bedroom: a History of Sex Research. (1994). Vern L. Bullough. New York: BasicBooks, 376 páginas.
Sex, Cells, and Same-Sex Desire: The Biology of Sexual Preference. (1995). John P. De Cecco & David Allen Parker, editores. New York: Harrington Park Press, 470 páginas.
Artículos.
Adolphs, R., Travel, D., Damasio, A.R. (1998). The human amygdala in social judgement. Nature, 393: 470-474
Allen, L.S., Gorski, R.A. (1990). Sex differences in the bed nucleus of the sria terminalis of the human brain. J.Comp.Neurol., 302: 697-706.
Allen, L.S., Hines, M., Shryne, J.E., Gorski, R.A. (1989). Two sexually dimorphic cell groups in the human brain. J.Neurosci., 9: 497-506.
Arendash, G.W., Gorski, R.A. (1983). Effects of discrete lesions of the sexually dimorphic nucleus of the preoptic area or other medial preoptic regions on the sexual behavior of male rats. Brain Research Bulletin, 10: 147-154.
Bakker, J., Van Ophemert, J., Slob, A.K. (1993a). Organization of partner preference and sexual behavior and its nocturnal rhythmicity in male rats. Behavioral Neuroscience, 107: 1049-1058.
Bakker, J., Van Ophemert, J., Slob, A.K. (1993b). Hormonal regulation of adult partner preference behavior in neonatally ATD-treated male rats. Behavioral Neuroscience, 107: 480-487.
Claro, F., Segovia, S., Guilamón, A., Del Abril, A. (1995). Lesions in the medial posterior region of the BST impair sexual behavior in sexually experienced and inexperienced male rats. Brain Research Bulletin, 36: 1-10.
Cohen, R.S. & Pfaff, D.W. (1992). Ventromedial hypothalamic neurons in the mediation of long-lasting effects of estrogen on lordosis behavior. Progress in Neurobiology, 38: 423-453.
Davis, P.G., McEwen, B.S., Pfaff, D.W. (1979). Localized behavioral effects of triated estradiol implants in the ventromedial hypothalamus of female rats. Endocrinology, 104: 898-903.
Decavel, C. & van den Pol, A (1990). GABA: a dominant neurotransmitter in the hypothalamus. J. Comp. Neurol., 302: 1019-1037.
Dulce-Madeira, M. & Lieberman, A.R. (1995). Sexual dimorphism in the mammalian lymbic system. Progress in Neurobiology, 45: 275-233.
Emery, D.A. & Sachs, B.D. (1976). Copulatory behavior in male rats with lesions in the bed nucleus of the stria terminalis. Physiology and Behavior, 17: 803-806.
Frankfurt, M., Gould, E., Woolley, C.S., McEwen, B.S. (1990). Gonadal steroids modify dendritic spine density in the ventromedial hypothalamic neurons: a Golgi study in the adult rat. Neuroendocrinology, 51: 530-535.
Gorski, R.A., Girdon, J.H., Shryne, J.E., Southman, A.M. (1978). Evidence for a morphological sex difference within the medial preoptic area of the rat brain. Brain Research, 148: 333-346
Hansen, S., Köhler, C., Glodstein, M., Steinbusch, H.V.M. (1982). Effects of ibotenic acid-induced neuronal degeneration in the medial preoptic area and the lateral hypothalamic area on sexual behavior in the rat. Brain Res., 239: 213-232.
Heimer, L. & Larsson, K. (1966/1967) Impairment of sexual behavior in male rats following lesions in the preoptic-anterior hypothalamic continuum. Brain Res., 3: 248-263.
Herbison, A.E. & Fénelon, V.S. (1995). Estrogen regulation of GABAA receptor subunit expression in preoptic area and bed nucleus of the stria terminalis of female rat brain. J. Neurosci., 15 (3): 2328-2337.
Hines, M., Allen, L.S., Gorski, R.A. (1992). Sex difference in subregions of the medial nucleus of the amygdala and the bed nucleus of the stria terminalis of the rat. Brain Research, 579: 321-326.
Jarry, H., Leonhardt, S., Wuttke, W. (1991). Gamma-aminobutyric acid neurons in the preoptic/anterior hypothalamic area synchronize the phasic activity of the gonadotropin-releasing hormone pulse generator in ovariectomized rats. Neuroendocrinology, 53: 262-267.
Jorge-Rivera, J.C. & Henderson, L.P. (1998). Androgenic steroids differentially modulate GABAA-mediated transmission in the VMN and the mPOA of the female rat brain. Soc.Neurosci.Abst. (en imprenta).
Jüptner, M. & Hiemke C. (1990). Sex differences in GABAA receptor binding in rat brain measured by an improved binding assay. Exp. Brain Res., 81: 297-307.
Kondo, Y., Sachs, B.D., Sakuma,Y. (1997). Importance of the medial amygdala in rat penile erection evoked by remote stimuli from estrous females. Behavioral Brain Research, 88 (2): 153-160.
Kruijver, F.P.M., De Jonge, F.H., Van den Broek, W.T., Van der Woude, T., Endert, E., Swaab, D.F. (1993). Lesions of the suprachiasmastic nucleus do not disturb sexual orientation of the adult male rat. Brain Research, 624: 342-346.
LeVay, S. (1991). A difference in hypothalamic structure between heterosexual and homosexual men. Science, 253: 1034-1037.
Liu, Y.-C., Salamone, J.D., Sachs, B.D. (1997). Lesions in medial preoptic area and bed nucleus of stria terminalis: differential effects on copulatory behavior and noncontact erection in male rats. J. Neuroscience. 17(13): 5245-5253.
Makara, G.B., Rappay, G., Stark, E. (1975). Autoradiographic localization of 3H-gamma-aminobutyric acid in the medial hypothalamus. Exp. Brain Res., 22: 449-455.
Masco, D.H. & Carrer, H.F. (1980). Sexual receptivity in female rats after lesion or stimulation in different amygdaloid nuclei. Physiology and Behavior, 24: 1073-1080.
McCarthy, M.M., Malik, K.F., Feder, H.H. (1990). Increased GABAergic transmission in medial hypothalamus facilitates lordosis but has the opposite effect in the preoptic area. Brain Research, 507: 40-45.
McCarthy, M.M., Masters, D.B., Fiber, J.M., López-Colomé, A.M., Beyer, C., Komisaruk, B.R., Feder, H.H. (1991). GABAergic control of receptivity in the female rat. Neuroendocrinology, 53: 473-479.
McCarthy, M.M. (1995) Functional significance of steroid modulation of GABAergic neurotransmission: analysis at the behavioral, cellular, and molecular levels. Hormones and Behavior, 29: 131-140.
Miller, L.D.P. & Aoki, A. (1991). Stereological analysis of the hypothalamic ventromedial nucleus. II. Hormone-induced changes in the synaptogenic pattern. Devl. Brain Res., 61: 189-196.
Morris, J.S., Öhman, A., Dolan, R.J. (1998). Concious and unconscious emotional learning in the human amygdala. Nature, 393: 467-470.
Nett, S.T., Jorge-Rivera, J.C., Myers, M., Clark, A.S., Henderson, L.P. (1998). Properties of GABAA receptors and sex-specific differences in benzodiazepine sensitivity of neurons expressing (-1 subunit mRNA in the preoptic area of the rat (sometido a publicación).
Nitecka, L. & Ben-Ari, Y. (1987). Distribution of GABA-like immunoreactivity in the rat amygdaloid complex. J. Comp. Neurol., 266: 45-55.
Nitecka, L. & Frotscher, M. (1989). Organization and synaptic interconnections of GABAergic and cholinergic elements in the rat amygdaloid nuclei: single and double-immunolabeling studies. J. Comp. Neurol., 279: 470-488.
Ogawa, S., Kow, L.M., Pfaff, D.W. (1991). Effects of GABA and related agents on the electrical activity of hypothalamic ventromedial nucleus in vitro. Exp. Brain Res., 85: 85-92.
Ondo, J., Mansky, T., Wuttke, W. (1982). In vivo GABA release from the medial preoptic area of diestrous and ovariectomized rats. Exp. Brain Res., 46: 69-72.
Oomura, Y., Aon, S., Koyama, Y., Fujita, I., Yoshimatsu, H. (1988). Central control of sexual behavior. Brain Research Bulletin, 20: 863-870.
Pérez, J., Zucchi, I., Maggi, A. (1986). Sexual dimorphism in the response of the GABAergic system to estrogen administration. J.Neurochem., 47: 1798-1803.
Pfaff, D.W. & Sakuma, Y. (1979a). Facilitation of the lordosis reflex of female rats from the ventromedial nucleus of the hypothalamus. J.Physiol., London, 288: 189-202.
Pfaff, D.W, & Sakuma, Y. (1979b). Deficit in the lordosis reflex of female rats caused by lesions in the ventromedial nucleus of the hypothalamus. J.Physiol., London, 288: 203-210.
Pfaff, D.W. & Schwartz-Giblin, S. (1988). Cellular mechanisms of female reproductive behaviors. En: The Physiology of Reproduction, editado por E.Knobil and J.Neil et al., Raven Press, Ltd., New York, 1487-1568.
Schumacher, M., Corini, H., McEwen, B.S. (1989). Regulation of high affinity GABAA-receptors in specific brain regions by ovarian hormones. Neuroendocrinology, 50: 315-320.
Smith, S.T,, Brenan, C.T., Clark, A.S., Henderson, L.P. (1996). GABAA receptor mediated-responses in the ventromedial nucleus of the hypothalamus of female and male rats. Neuroendicrinology, 64: 103-113.
Sördersten, P., Hansen, S., Srebro, B. (1981). Suprachiasmatic lesions disrupt the daily rhythmicity in the sexual behavior of normal male rats and male rats treated neonatally with antiestrogen. J.Endocrinology, 88:125-130.
Sundström, I., Ashbrook, D., Backström, T (1997). Reduced benzodiazepine sensitivity in patients with premenstrual syndrome: a pilot study. Psychoneuroendocrinology, 22 (1), 25-38.
Swaab, D.F., Fliers, E., Partiman, T. (1985). The suprachiasmatic nucleus of the human brain in relation to sex, age and dementia. Brain Research, 342: 37-44.
Swaab, D.F. & Hofman, M.A. (1988). Sexual differentiation of the human hypothalamus: ontogeny of the sexually dimorphic nucleus of the preoptic area. Dev. Brain Res., 44: 314-318.
Swaab, D.F. & Hofman, M.A. (1990). An enlarged suprachiasmatic nucleus in homosexual men. Brain Research, 537: 141-148.
Swaab, D.F. & Hofman, M.A. (1995). Sexual differentiation of the human hypothalamus in relation to gender and sexual orientation. Trends in Neuroscience, 18: 264-270.
Swaab, D.F., Slob, A.K., Houtsmuller, E.J., Brand. T., Zhou, J.N. (1995). Increased number of vasopressin neurons in the suprachiasmatic nucleus (SCN) of bisexual adult male rats following perinatal treatment with the aromatase blocker ATD. Developmental Brain Research, 85: 273-279.
Szechtman, H., Caggiula, A.R., Wulkman, D. (1978). Preoptic knife cuts and sexual behavior in male rats. Brain Research, 150: 569-591.
Wagner, S., Castel, M., Gainer, H., Yarom, Y. (1997). GABA in the mammalian suprachiasmatic nucleus and its role in diurnal rhythmicity. Nature, 387 (5): 598-603.
Weller, K.L. & Smith, D.A. (1982). Afferent connections to the bed nucleus of the stria terminalis. Brain Research, 232: 255-270.
Wisden, W., Laurie, D.J,, Monyer, H., Seeburg, P.H. (1992). The distribution of 13 GABAA receptor subunit mRNAs in the rat brain. I. Telencephalon, diencephalon, mesencephalon. J.Neurosci., 12: 1040-1062.
Young, L.J., Wang, Z., Insel, T.R. (1998). Neuroendocrine bases of monogamy. Trends Neurosci., 21, 71-75.
Zhou, J.N., Hofman, M.A., Gooren, L.J.G., Swaab, D.F. (1995). A sex difference in the human brain and its relation to transsexuality. Nature, 378: 2, 68-70.
Punteros de interés
