
MICROBIOLOGIA |
Métodos de estudio de los cambios estructurales en ecosistemas microbianos edáficos y su aplicación ambiental |
|
©
Leonidas Carrasco Letelier
,
2003
lcarrasco@netexplora.cl |
RESUMEN[ABSTRACT]
En la última década se han incrementado los estudios de la microflora de suelos debido a que se ha reconocido su rol fundamental en el soporte de las funciones y servicios ecosistémicos. Por esto, el análisis de la estructura de estos ecosistemas permitiría el monitoreo y predicción de los cambios en la calidad del suelo. Sin embargo, este análisis estructural no ha podido llevarse a cabo por la falta de información sobre la biodiversidad de estos ecosistemas, especialmente, luego de que Torsvik y colaboradores (1980, 1990) constataran la ineficiencia y poca representatividad del método microbiológico tradicional. En estos últimos años se han desarrollado métodos nuevos para el estudio de los cambios estructurales de los ecosistemas microbianos edáficos que, si bien se han desarrollado para la evaluación de impactos ambientales, su uso en ésta área ha sido reducido. Este artículo revisa las ventajas y desventajas de los principales métodos que hoy existen; analizados desde las necesidades de un país en vías de desarrollo que requiere evaluar sus impactos ambientales sobre el suelo y relevar su biodiversidad microbiana, como recurso biotecnológico para el país.
De los compartimientos ambientales principales, el suelo es el más complejo de estudiar y analizar, debido a su complejidad estructural y carencia de métodos eficientes para su evaluación, especialmente su componente biológica. Además, presenta los mayores desafíos científicos, en términos ambientales, dada la gran cantidad de servicios y funciones ecosistémicas que presta a la Humanidad (descomposición de material vegetal y animal, ciclos biogeoquímicos, sumidero de carbono, soporte para la producción vegetal y animal, etc.) (Turco et al., 1994; Widmer et al., 2001). Por lo que, actualmente, un elemento central del estudio de los suelos es la evaluación de los cambios de su calidad, donde la componente biológica ha mostrado ser de gran relevancia. Especialmente la flora microbiana (Stenberg, 1997; Stenberg, 1999), ya que es la encargada de sustentar las diferentes funciones del suelo, por lo cual una alteración de ésta - por cambios de cobertura vegetal u otro tipo de impacto ambiental - puede desencadenar cambios drásticos en las propiedades globales del suelo, como su fertilidad (Baath et al., 1998; Bossio et al., 1998; Di Giovanni et al., 1999; Haldeman et al., 1995; Ibekwe Abasiofiok & Kennedy, 1998; Ovreas et al., 1998; Rundgren et al., 1998; Zogg et al., 1997).
A pesar de estos antecedentes, los estudios microbiológicos del suelo
fueron a menudo subvalorados, principalmente por ser engorrosos y complejos
(Turco et al., 1994; Widmer et al., 2001); lo cual, los dejó en desventaja
respecto a los análisis de parámetros fisicoquímicos (contenido
de arcilla, limo y arena; acidez; capacidad de retención de agua, capacidad
de intercambio catiónico, carbono y nitrógeno total, etc.). Sin
embargo, éstos últimos no han permitido explicar los cambios generados
por los diferentes tipos de impactos ambientales en la calidad del suelo (Turco
et al., 1994; Widmer et al., 2001); ya que, los cambios de la calidad responden
principalmente a los cambios de su componente biológica (Dick, 1994).
Un análisis más profundo de ésta componente del suelo se
ha intentado realizar a través de la evaluación de las propiedades
funcionales del ecosistema microbiano, mediante la determinación de:
la biomasa microbiana, tasa de respiración microbiana, la tasa de transformación
de nutrientes específicos (actividades enzimáticas generales y
específicas), y parámetros ecofisiológicos (Insam &
Öhlinger, 1995). Y aunque, este tipo de estudios logró mostrar su
validez en el monitoreo de los cambios de la calidad del suelo, no ha permitido
develar el funcionamiento interno del ecosistema microbiano edáfico,
manteniendo a éste en una condición de "caja negra".
Lo cual, impide comprender cómo los diferentes impactos ambientales afectan
la fragilidad y sustentabilidad de estos ecosistemas (Insam, 2001; Tiedje et
al., 1999). Por esta razón, en la última década, en diferentes
trabajos, se ha planteado como prioritario el estudio de los cambios provocados
por los impactos ambientales en la estructura de los ecosistemas microbianos
edáficos; de manera de poder evaluar y predecir los cambios futuros de
la calidad de los suelos, basados en los cambios de características del
ecosistema (como cambios en la riqueza y equitatividad) y en el hallazgo de
bioindicadores y biomarcadores (Rundgren et al., 1998; Schutter & Dick 2000;Tiedje
et al., 1999; Turco et al., 1994).
El principal motivo que obstaculizó un mayor avance de los estudios de la microflora del suelo fue el empleo del método microbiológico clásico, que era muy lento y engorroso; debido al gran número de etapas requeridas para lograr el aislamiento e identificación taxonómica de los diferentes organismos (Turco et al., 1994). Esta metodología se empleo hasta mediados de los años 80, cuando el desarrollo de estudios de biología molecular de la microflora del suelo constataron que el método clásico había evaluado menos del 1% de los microorganismos (Tiedje et al., 1999; Torsvik, 1980; Torsvik et al., 1990; Turco et al., 1994). Estos hallazgos, conjuntamente con el auge de la biotecnología y nuevas metodologías de la biología molecular - como la amplificación de ADN con PCR -, conllevó al desarrollo de nuevas áreas de la ciencia, como la Ecología Molecular Microbiana (Rosado et al., 1997), que incrementaron los estudios taxonómicos de la microflora del suelo (Insam, 2001; Tiedje et al., 1999; Turco et al., 1994; Zelles, 1999). Desafortunadamente, la mayor parte de estos estudios no han logrado apartarse de los objetivos biotecnológicos, existiendo un avance pobre en entender los mecanismos internos de la "caja negra", y sus relaciones con la calidad del suelo y los impactos ambientales.
Este ensayo tiene por objetivo revisar y analizar las nuevas metodologías desarrolladas para el estudio de los cambios estructurales de los ecosistemas microbianos de suelos, así como sus potencialidades en estudios con objetivos ambientales, tanto para el diagnóstico como para la predicción de los cambios de la calidad de los mismos.
Estudios por métodos tradicionales
El estudio del suelo es un problema complejo, debido a dos grandes causas: primero por ser de una gran complejidad estructural, ya que presenta fases sólidas, líquidas y gaseosas, más sus respectivas interfases; y segundo, por la carencia de métodos de análisis desarrollados para evaluar adecuadamente las bases biológicas que sustentan sus funciones y servicios (Turco et al., 1994).
Entre los métodos empleados para el estudio de los suelos, los análisis físicos son los de mayor eficiencia y confiabilidad, dado que fueron específicamente desarrollados para el análisis de este compartimiento ambiental. En un grado intermedio de confiabilidad, se encuentran los análisis químicos del suelo, donde los principios de la química de sistemas acuosos son validos, en la mayor parte de los casos, si se logra obtener soluciones homogéneas del analito buscado. En otros casos, como la determinación de la acidez, el suelo actúa como un coloide, por lo cual dependiendo de la distancia entre el electrodo y el sedimento se obtendrán valores diferentes, afectando la reproducibilidad del método (McLean, 1982). Por estas razones, en la década del 60, cuando se realizaron los relevamientos de suelos del mundo, se emplearon esencialmente métodos físicoquímicos (Falkowski et al., 2000; Scholes et al., 1995).
El método microbiológico clásico que se empleo en el estudio del suelo, originalmente desarrollado para muestras acuosas, consta de cinco etapas: (i) dilución de la muestra (paso optativo); (ii) cultivo en medios de cultivo generales, que idealmente proporcionan las condiciones de crecimiento para todos los microorganismos de la muestra; (iii) cultivo en medios específicos, para la selección de grupos de organismos; (iv) conteo; e (v) identificación taxonómica, mediante la evaluación de parámetros bioquímicos, como la fermentación de diferentes substratos (Parkinson et al., 1971; Schinner et al., 1996). A pesar de ser una metodología eficiente en estudios de microbiología humana y de sistemas acuáticos, su empleo en el estudio de suelos se realizó con ciertos reparos, ya que se asumía que sus premisas no se cumplían totalmente. Primero, porque el supuesto de una dilución homogénea de la muestra en el caso de los suelos es irreal, ya que nunca se logra alcanzar dicha condición; segundo, debido a que los organismos están estrechamente unidos a las partículas del suelo, no se puede lograr una dilución representativa de todas las especies; y tercero, las condiciones del medio de cultivo general, no aseguran el crecimiento de todos los microorganismos presentes en la muestra de suelo, ya que las condiciones aeróbicas, así como las concentraciones de los substratos, no serán optimas para todas las comunidades microbianas. Por lo cual, se aceptaba que la metodología clásica sólo lograba la identificación de una submuestra "representativa" de los organismos aeróbicos presentes en las muestras analizadas (Rosado et al., 1997; Turco et al., 1994; Van Elsas et al., 1998; Zelles, 1999). Con los hallazgos de Torsvik et al. (1980, 1991), que señalaban que entre el 99,5 a 99,9 % de los microorganismos de suelo no eran cultivables por el método microbiológico clásico, se estableció la poca representatividad estadística del método clásico. A lo que se sumaron posteriormente, las discordancias entre las clasificaciones taxonómicas realizados mediante el método clásico y los estudios del ADN (Jarvis & Tighe, 1994).
Nuevas metodologías
La creciente preocupación por desarrollar bioindicadores para monitorear los cambios en la calidad del suelo, provocados por: derrames y disposición de xenobióticos, contaminación por plaguicidas, procesos de erosión o el incremento periódico del uso de fertilizantes para lograr mantener las producciones agrícolas; hicieron que la investigación de suelos se volviese prioritaria, y que la mayor parte de la investigación se haya realizado tan sólo en la pasada década (Insam, 2001), especialmente sobre los cambios estructurales de los ecosistemas microbianos edáficos vinculados impactos ambientales (Baath et al., 1998; Bossio et al., 1998; Di Giovanni et al., 1999; Haldeman et al., 1995; Ibekwe Abasiofiok & Kennedy, 1998; Ovreas et al., 1998; Zogg et al., 1997). Aunque mediante el análisis funcional del ecosistema microbiano edáfico se logró definir relaciones con los cambios en la calidad del suelo; estos hallazgos son difíciles de interpretar en términos ecosistémicos (Schinner et al., 1996), por lo cual, su interpretación y uso se ha realizado en forma reduccionista. Un ejemplo de esto es la evaluación de la fertilidad del suelo basada en relación carbono nitrógeno (Schinner et al., 1996), lo cual no explicaba las pérdidas de productividad e incrementos en los requerimientos de fertilizantes; situación que sólo fue criticado desde al ambientalismo, especialmente por visiones holísticas como la Agroecología, que siempre insistieron en una evaluación de los suelo como un ecosistema y no como un simple soporte inerte (Altieri, 1999).
Los nuevos estudios de la estructura del ecosistema microbiano han generado información importante para la comprensión de los mecanismos que dan origen a las funciones y servicios de la "caja negra". Lamentablemente, la mayor parte de estos estudios se ha centrado en casos de contaminación química (Rosado et al., 1997; Zelles, 1999), y últimamente en la evaluación de los efectos de plantas transgénicas (Germida et al., 1998a; Germida et al., 1998b; Siciliano et al., 1998; Siciliano & Germida, 1999; Tiedje et al., 1999); reduciendo la potencialidad de las nuevas herramientas, al circunscribirlas en la búsqueda o monitoreo de organismos degradadores de los contaminantes, debido a la alta influencia de intereses económicos sobre nuevos recursos biotecnológicos; por esto, la evaluación de los índices de diversidad u otros descriptores ecológicos de las comunidades y el efecto de los impactos antrópicos sobre ellas ha sido escaso, exceptuando estudios de Degens (Sparling & Degens, 1999; Degens & Vojvodic-Vucovik, 1999) y Zelles (1999).
Actualmente existe una gran variedad de estudios sobre microorganismos del suelo, que se pueden agrupar en cinco grandes áreas: (i) reclasificación taxonómica (Graham et al., 1995; Jarvis & Tighe, 1994); (ii) monitoreo de genes en el ambiente (Van Elsas et al., 1998); (iii) monitoreo de especies transgénicas (Siciliano et al., 1998); (iv) relevamiento de microorganismos de interés biotecnológico; y (v) estudios de carácter ambiental (Maire et al., 1999; Van Elsas et al., 1998). Para estos propósitos se han desarrollado cuatro diferentes metodologías: (a) análisis de uso de substratos in vitro, (b) análisis del uso de substratos in situ, (c) análisis de patrones de ácidos grasos y (d) análisis de patrones de ADN (Insam, 2001).
Análisis del uso de substratos in vitro
El análisis de patrones de uso de substratos a nivel comunitario (abreviado en inglés como CLSU fingerprint [1], de la expresión "community level substrate utilization fingerprints") consiste en analizar el comportamiento de las comunidades microbianas del suelo en 96 medios de cultivo diferentes en forma simultánea (dispuestos en una placa de microtitulación). En si es una variación del método microbiológico clásico, introduciendo como innovación el uso de medios de cultivo diferentes, cada uno con una substancia indicadora coloreada, que señala el tipo y grado de actividad realizada en cada medio de cultivo. Este método se puede emplear tanto para especies bacterianas aisladas como para comunidades completas. Esta técnica está patentada bajo el nombre de BIOLOG, empresa que comercializa dichas placas de microtitulación. En si el método es muy simple, y permite identificar diferencias en la reactividad con los substratos, ya que se puede evaluar colorimétricamente (Siciliano et al., 1998; Waldrop et al., 2000; Widmer et al., 2001).
La característica más relevante de este método es permitir identificar y cuantificar un patrón metabólico, en forma rápida y poco costosa, que permite el relevamiento de grandes cantidades de muestras y el agrupamiento de suelos, en función de los patrones encontrados (Wildmer et al., 2001). Esta técnica ha demostrado su capacidad de realizar identificaciones taxonómicas, ya que existen patrones de reacción definidos por especie. A pesar de estas ventajas, el método presenta problemas similares al método clásico, ya que: utiliza una dilución del suelo; sus condiciones de cultivo sólo detectan actividades de algunos microorganismos aeróbicos de la muestra, con lo cual su representatividad estadística es cuestionables (Insam, 2001). Además, a pesar que esta metodología se denomina patrón de uso de substratos, revela poco sobre las alteraciones ocurridas en el ámbito fisiológico o estructural en el ecosistema microbiano, porque en el suelo existen actividades catabólicas redundantes, es decir, sustentadas por más de un tipo de microorganismo, de esta forma podrían existir reducciones en la riqueza de los ecosistemas y no ser detectadas a través de esta metodología (Dick, 1994; Dick et al., 2000); asimismo, las actividades de los consorcios microbianos, realizados para la complementación de los metabolismos microbianos, no pueden detectarse, ya sea su desagregación física (en la etapa de dilución) o porque las condiciones son restrictivas para algunos de los integrantes del consorcio.
Análisis del uso de substratos in situ
Como solución a los problemas de del análisis del patrón de uso de substratos in vitro (BIOLOG), un grupo de investigación de Nueva Zelandia, creó una forma optimizada de éste, mediante su evaluación in situ (Degens, 1999). Éste análisis consiste en agregar al suelo estudiado una mezcla de substratos conocidos que posteriormente es recuperada y evaluada. De esta manera, se puede determinar cual ha sido la catabolización de cada substrato en el suelo - lo que se ha denominado diversidad funcional catabólica - . Esta estrategia de evaluación de la diversidad es muy interesante, aunque más compleja y costosa que BIOLOG, y quizás esto explique que el único país donde se utiliza es Nueva Zelandia, lugar donde se creo esta metodología. La desventaja principal de este método es no permitir evaluar los cambios en la estructura del ecosistema microbiano en términos taxonómicos, ya que no es posible asignar las funciones catabólicas a grupos microbianos específicos (Degens & Sparling, 1996; Degens & Harris, 1997; Degens 1998a; Degens 1998b; Degens, 1999; Degens & Vojvodic-Vucovik, 1999; Degens et al., 2000; Degens et al., 2001). Además existe la posibilidad de interferencia en los resultados por la actividad catalítica de las sustancias húmicas (Klavins et al., 2001) o enzimas libres del suelo [2] (Dick, 1994).
Análisis de patrones de ácidos grasos
El análisis de ácidos grasos se basa en el estudio de los ácidos grasos, componentes esenciales de la membrana celular de las células vivas, que en condiciones naturales son una proporción relativamente constante de la biomasa de los organismos (Lechevalier, 1977). De esta manera, es posible obtener un patrón representativo de especies aisladas o de ecosistema microbiano enteros (Zelles, 1999), ya que sólo las células vivas viables poseen membranas no alteradas, con ácidos grasos como los fosfolípidos (denominados en la literatura inglesa como PLFA [3]). El análisis del patrón de éstos ácidos grasos, que se realiza a través de la identificación y cuantificación de sus ésteres metílicos (denominados en la literatura inglesa como FAME [4]), mediante cromatografía gaseosa o de capa fina; cuyo patrón es característico para cada género y especie (Liu, 1994; MIDI, 1990; Turco et al., 1994). Este tipo de análisis de los ecosistemas microbianos se ha utilizado con tres variantes: (i) aplicación a microorganismos aislados mediante técnicas microbiológicas tradicionales (métodos FAME-MIDI) (Figura 1), donde la identificación se realiza mediante el empleo de una base de datos (como la MIS [5]) (MIDI, 1990); (ii)identificación de relaciones entre grupos de microorganismos basado en ácidos grasos característicos de especies o géneros, como el caso de las evaluaciones de ergosterol (ácido graso propio de hongos) (Grierson & Adams, 2000; Joergensen, 2000; Kato & Wickner, 2001; Ruzicka et al., 2000); y (iii) el análisis de ecosistemas microbianos enteros, sin etapas de aislación de especies, donde se estudian ácidos grasos que identifiquen comunidades u organismos específicos, con fines ambientales (Tabla 1 y Figura 2) (Jansa et al., 1999; Nielsen & Petersen, 2000; Olsson et al., 1999; Olsson & Persson, 1999; Schutter & Dick, 2000). Ésta última estrategia permite definir y agrupar suelos con las características funcionales determinadas (niveles de carbono, fertilidad, actividades enzimáticas, etc.) (Maire et al., 1999), así como la identificación de las comunidades de microorganismos (Zelles, 1999).
Tabla 1. Resumen de los patrones mas importantes de fosfolípidos que pueden ser utilizados para determinar la composición comunitaria de la microbiota del suelo (Zelles, 1999).
|
Grupo microbiano
|
Patrón de fosfolípidos
|
| Archaebacteria | Residuos grasos con una unión éter a glicerol |
| Bacterias Anaeróbicas | Contienen esfingolípidos que están ausentes en los aeróbicos |
| Bacterias en general | Ácidos grasos saturados y monoinsaturados con unión éster a glicerol |
| Bacterias gram negativa | Contienen mayor cantidad de ácidos grasos hidroxilados |
| Bacterias gram positiva | Contienen mayor cantidad de ácidos grasos ramificados. |
| Cyanobacteria(y eucariotas) | Contienen ácidos grasos poli-insaturados. |
| Hongos | Presentan un fosfolípidos específico, como: 18:2 6 ó ergosterol. |
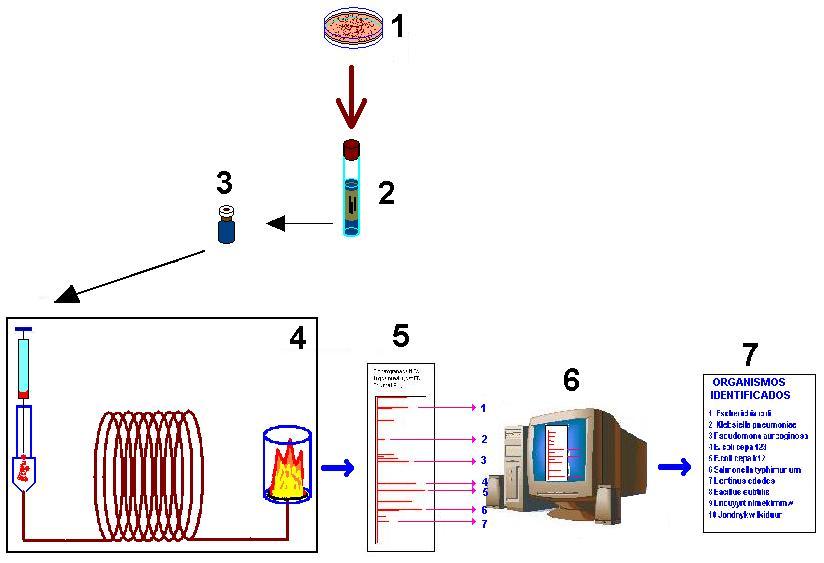
Figura 1. Procedimiento general seguido por el método de análisis FAME: (1) cultivo microbiológico puro; (2) saponificación / metilación; (3) extracción de ésteres metílicos; (4) análisis por cromatografía gaseosa con detector FID; (5) etapa de integración; (6) denominación de señales y comparación del patrón con una base de datos; (7) Identificación de la especie microbiana (MIDI, 1990).
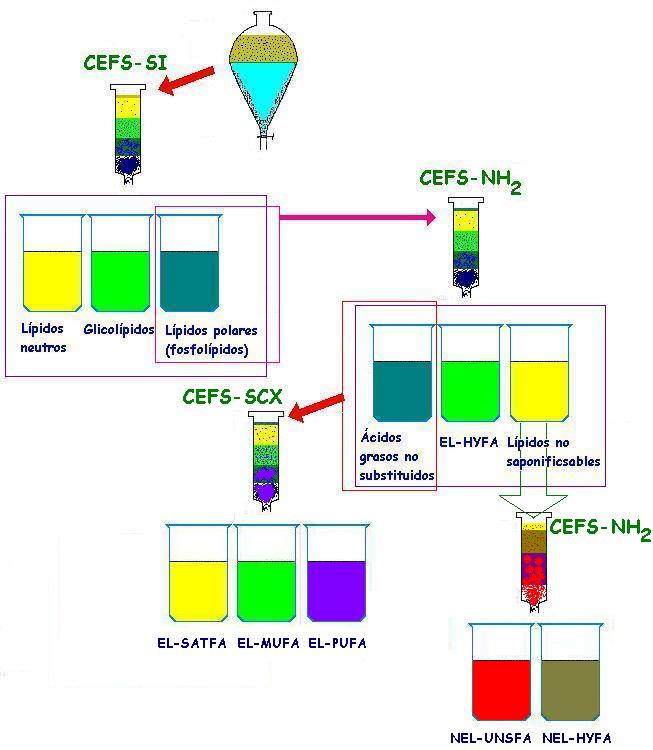
Figura 2. Diferentes etapas de extracción y separación de los fosfolípidos, mediante columnas de extracción en fase sólida (CEFS) con grupos de: sílica (SI), aminopropil (NH2) y ácido bencensulfónico (SCX). Para separar ácidos grasos hidroxilados (EL-HYFA), saturados (EL-SATFA), monosaturados (EL-MUFA), poliinsaturados (EL-PUFA) y de ácidos grasos no esterificados no hidroxilados (NEL-UNSFA) e hidroxilados (NEL-HYFA) (Zelles, 1999).
Análisis de patrones de ADN
El desarrollo de las técnicas para el análisis de patrones ADN ha ido de la mano con la evaluación del riesgo asociado a la liberación (deliberada o involuntaria) de organismos transgénicos al medio ambiente. Las diferentes técnicas presentan básicamente cuatro etapas fundamentales: (i) aislamiento de los ácidos nucleicos (ADN y/o ARN) microbianos del suelo; (ii) amplificación, mediante PCR; (iii) análisis de los patrones de restricción; y (iv) cuantificación. Además, como etapa complementaria está la posibilidad del clonamiento del material aislado para la identificación de sus funciones, dependiendo de los objetivos de la investigación (Tiedge et al., 1999). Debido a la gran cantidad de las variantes de cada etapa, cada una con una denominación propia (Figura. 3), que explicarlas en detalle está fuera de los objetivos de este artículo, por lo cual se recomienda revisar los trabajos del grupo de van Elsas (van Elsas et al., 1998; Rosado et al., 1997), donde se han resumido la mayor parte de todas las variantes conocidas de esta metodología.
Sobre este tipo de análisis, Insam (2001) ha argumentado, que debido a sus capacidades taxonómicas en el futuro reemplazarán a los análisis de patrones de ésteres metílicos de ácidos grasos. Esto porque los estudios de ácidos nucleicos pueden asociar directamente la información encontrada con genotecas microbianas, permitiendo la identificación a nivel de género y especie; lo que permitiría la clasificación taxonómica en casos de especies desconocidas. Sin embargo, esta metodología sólo ha demostrado su eficiencia en sistemas simplificados, donde las interferencias en la detección de genes (o grupos de genes) específicos han sido eliminadas mediante la amplificación por PCR con sondas específicas para el material buscado (Germida et al., 1998a; Germida et al., 1998b; Siciliano et al., 1998; Siciliano & Germida, 1999). Pero, en el análisis de muestras de suelo crudas, sin aislamiento previo de las especies, surgen problemas serios para que se mantengan los niveles de eficiencia logrados con genes y microorganismos aislados (Harry et al., 1999; Harry et al., 2000; Harry et al., 2001; Roose-Amsaleg et al., 2001). Primero, porque en la etapa de extracción del ADN o ARN desde el suelo se co-extraen sustancias húmicas (dada sus similitudes estructurales y fisicoquímicas), que inhiben la actividad de la PCR reduciendo la eficiencia de la amplificación. Segundo, el ADN extraído del suelo es una mezcla de ADN genómicos, por lo que existirá competencia favorable por los primers [6] para aquellos genomas que dominen en ecosistema, en la etapa de amplificación con PCR. Esto plantea el riesgo de no poder amplificar ni detectar el ADN de especies poco representadas en el ecosistema, ni de poder asignar los fingerprints de ADN del suelo a especies microbianas específicas. Por lo tanto, las aplicaciones ambientales de este tipo de técnicas presentarán resultados potenciales muy similares a los actuales análisis de patrones de ésteres metílicos de ácidos grasos de muestras de suelo entero.
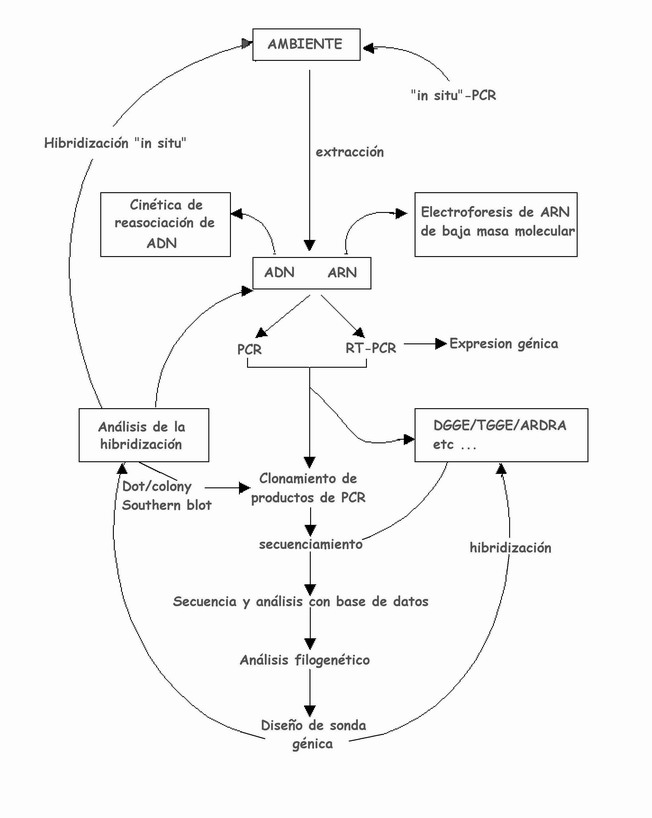
Figura 3. Diagrama esquemático que muestra en general las diferentes técnicas de biología molecular empleadas para la caracterización de microorganismos de muestras ambientales (Rosado et al., 1997).
Aplicaciones ambientales
El estudio de las comunidades microbianas con objetivos ambientales se ha centrado en el análisis de efectos del cambio de cobertura vegetal, análisis de la rizósfera, evaluación del estrés ambiental causado por xenobióticos, y en la optimización de procedimientos para el análisis de microorganismos del suelo, sin etapas intermedias de cultivo.
El análisis de patrones de sustrato in situ se ha empleado para evaluar: efecto de enmiendas (Degens, 1998b); modulaciones de la diversidad catabólica provocadas por los niveles de carbono orgánico del suelo (Degens et al., 2000); y cambios en la diversidad funcional y su fragilidad en diferentes coberturas vegetales (bosque nativo, pasturas, cultivos y plantaciones de pino) (Degens & Vojvodic-Vukovic, 1999; Sparling & Degens, 2000); En esta última aplicación se destaca el cálculo de índices de riqueza y equitatividad, basados en la diversidad funcional catabólica; con lo cual se ha intentado evaluar los cambios en fragilidad del ecosistema edáfico frente a nuevos impactos ambientales. Sin embargo, este análisis de la diversidad catabólica, aun no ha logrado mostrar su correlación con análisis taxonómicos, específicamente con el PLFA (Degens 1998a; Wildmer et al., 2001).
Por otra parte, los análisis de patrones de ésteres metílicos de ácidos grasos han mostrado su mayor amplitud en sus aplicaciones ambientales, utilizándose en casi todas las áreas mencionadas al comienzo de esta sección. Se han empleados con éxito en el análisis de cambios de cobertura vegetal, como bosque tropical por tierras de cultivo (Waldrop et al., 2000), para evaluar diferentes tipos de cultivo agrícolas (Bossio et al., 1998; Maire et al., 1999; Zelles et al., 1992) y sus relaciones con propiedades funcionales, especialmente actividades enzimáticas (Grierson & Adams, 2000, Kandeler et al., 1998; Stenberg, 1997; Stenberg, 1999; Sundh et al., 2000). Asimismo se han ocupado en el estudio del funcionamiento de la rizósfera (Jansa et al., 1999; Joergensen, 2000; Olsson & Persson, 1999; Olsson et al., 1999) y las alteraciones provocadas, en este contexto, por especies transgénicas (Germida et al., 1998a; Germida et al., 1998b; Siciliano & Germida, 1999; Siciliano et al., 1998). En el estudio de xenobióticos, las evaluaciones se han focalizado en el análisis de los efectos de la contaminación por cadmio, cobre, níquel, plomo y zinc (Baath et al., 1998; Frostegard et al., 1993). Donde se ha logrado definir un patrón especifico de PLFA, para este tipo de contaminación, que está directamente relacionado con el grado de contaminación (Zelles, 1999).
Por otra parte, el análisis de patrones de ácidos nucleicos se ha empleado hasta el momento en el análisis taxonómico de comunidades relacionadas a contaminaciones específicas por metales (Brim et al., 1999) y pesticidas (Ka et al.,1994, Tiedge et al., 1999). Sin existir aun una proyección ambiental de este tipo de estudios, ya que son demasiado reduccionistas; por lo cual, la mayor parte de las publicaciones de esta metodología tratan sobre optimizaciones de las técnicas (Insam, 2001; Krsek & Wellington, 1999; Rosado et al., 1997; Tiedge et al., 1999; van Elsas et al., 1998).
En general, analizando los patrones dados por los resultados de BIOLOG, PLFA
y de ADN, empleados en forma simultánea para un mismo tipo de suelo,
Wildmer et al. (2001) demostraron que los resultados, de cada tipo de análisis,
eran concordantes, ya que generaban un resultado similar en el agrupamiento
de los suelos. Sin embargo, al realizar el análisis de la estructura
del ecosistema edáfico, mediante un análisis de clusters de los
resultados obtenidos con cada método, los agrupamientos dados por cada
método no fueron similares. Esto demostró que los métodos,
si bien, evalúan aparentemente la misma variable del suelo; no lo hacen
de la misma forma.
Conclusiones
El estudio de los cambios estructurales en el ecosistema microbiano, desde un punto de vista ambiental, tiene como objetivos: evaluar la calidad del suelo y definir bioindicadores y/o biomarcadores. A esto se debe agregar, que estos estudios deben permitir tanto la evaluación del estado actual de los ecosistemas edáficos, como permitir predecir los cambios que se provocarían en diferentes escalas de trabajo, única forma de poder integrar esta información con otros antecedentes del problema ambiental. En este contexto, aunque todos los métodos revisados han sido planteados y usados con fines ambientales, se destacan entre ellos el análisis de patrones de ésteres metílicos de ácidos grasos y de patrones de ácidos nucleicos; ya que, en términos taxonómicos permitirían comprender los cambios estructurales y simultáneamente realizar un relevamiento de la biodiversidad, información de utilidad para la restauración o evaluación de suelos con características edafológicas similares. Por otro lado, los estudios realizados mediante la evaluación de utilización de substratos, aunque generan resultados aceptables, en términos agronómicos, para el agrupamiento de suelos; no lo son en términos ambientales. Esto, porque la información que proporcionan no dejan de ser un análisis elegante de las propiedades funcionales de la "caja negra". Lo único rescatable de este último tipo de análisis son las tentativas del grupo de Nueva Zelandia (Degens & Vojvodic-Vukovic, 1999) para calcular parámetros ecológicos (riqueza y equitatividad), situación que no se ha dado en el resto de los métodos.
En el actual escenario de trabajo que dominan el análisis de los cambios en la calidad de los suelos, provocados por los diferentes impactos ambientales, donde no existe información de la biodiversidad microbiana de los suelos; los métodos a emplear requieren tanto evaluar los cambios de la calidad como proporcionar información de base sobre los suelos dominantes de cada país. Lo cual, plantea una gran exigencia a la evaluación ambiental de la calidad de los suelos, por la escasez de investigadores y laboratorios capaces de llevar a cabo esta tarea, especialmente en América Latina.
Por esta razón, es que el análisis de los patrones de ésteres de ácidos grasos se presenta con ventaja sobre los análisis de ácidos nucleicos, ya que: generan resultados similares en el análisis de muestras crudas de suelo; y el costo del análisis de ácidos nucleicos, en términos de infraestructura y personal capacitado puede llegar a ser prohibitivo en países con bajo presupuesto de investigación. Además, el problema de la interferencia de las sustancias húmicas que se incrementa en suelos de interés agrícola, por su mayor contenido de materia orgánica; hacen prudente no utilizar estos métodos hasta que se terminen de estandarizar y determinar las condiciones óptimas de trabajo.
Por su parte, los análisis de FAME, plantean como otra ventaja la posibilidad de realizar inferencias a escalas de trabajo superiores a las microbiológicas, sin una identificación taxonómica. Un ejemplo de esto, es que la definición de un incremento del dominio de las poblaciones de hongos en un ecosistema (situación de suelos forestados), permite asumir eventos de "water repellency" [8] que podrían provocar cambios en la geomorfología del paisaje por: disminución de la tasa de percolación y recarga de acuíferos; incremento de la escorrentía y, de esta manera, de las tasas de erosión.
En resumen, en países latinoamericanos cuya biodiversidad microbiana se desconoce, así como las alteraciones provocadas por los impactos ambientales, y en un escenario de trabajo con recursos humanos e infraestructura limitada; los estudios de la calidad del suelo, a través de la evaluación de las alteraciones en la estructura de los ecosistemas microbianos, deberían iniciarse en base a análisis de FAME. Los que en el futuro deberían ser complementados con el análisis de patrones de ácidos nucleicos, con el objeto de registrar dicha biodiversidad, en términos taxonómicos. Porque en un futuro no muy lejano esta será una nueva forma de valoración económica de los suelo, dada su potencialidad en recursos biotecnológicos. Los cuales sólo podrán ser capitalizados por cada país si son estudiados en un contexto nacional, y no por empresas transnacionales.
Notas
[1] Fingerprints: es un término inglés cuya traducción
es huella digital, pero que la biología molecular lo utiliza con el significado
de patrón de identificación.
[2] En el suelo existe la posibilidad de encontrar enzimas estabilizadas por
su unión a la arcilla o las sustancias húmicas, que no corresponden
neo-síntesis de células, que se denominan enzimas libres.
[3] PLFA: phospholipids fatty acids, expresión en inglés que significa,
ácido graso fosfolipídico.
[4] FAME: fatty acids methyl esters, expresión en inglés que significa
ésteres metílicos de ácidos grasos.
[5] MIS: microbial identification system, perteneciente a la empresa MIDI.
[6] Primers: secuencias cortas de ácidos nucleicos, que complementan
trozos específicos en el ADN, y que permiten el enganche e inicio de
la copia de la hebra complementaria del ADN que se desea amplificar por PCR.
[7] Water repellency: es un estado, en el cual el suelo presenta un comportamiento
hidrofóbico.
Bibliografía
Altieri, M (1999). Agroecología: bases científicas para una agricultura sustentable. Editorial Nordan Comunidad, Montevideo, Uruguay, 338 pp.
Baath, E, Diaz Ravina, M, Frostegard, A, Campbell, A & Collin, D (1998). Effect of metal-rich sludge amendments on the soil microbial community. Applied Environmental Microbiolology., 64: 238-245.
Bossio, DA, Scow, KM, Gunapala, N & Graham, KJ (1998). Determinants of soil microbial communities: effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microbial Ecology., 36: 1-12.
Brim, H, Heuer, H, Krögerrecklenfort, E, Mergeay, M & Smalla, K (1999). Characterization of the bacterial community of a zinc-polluted soil. Can. J. Microbiol., 45: 326-338.
Ka, JO, Holben, WE & Tiedje, JM (1994). Genetic and phenotypic diversity of 2,4-dichlorophenoxyacetic acid (2,4-D)-degrading bacteria isolated from 2,4-D treated field soils. Applied Environmental Microbiology., 60: 1106 - 1115.
Degens, BP & Harris, JA (1997). Development of a physiological approach to measuring the catabolic diversity of soil microbial communities. Soil Biology and Biochemistry., 29: 1309-1320.
Degens, BP & Sparling, GP (1996). Changes in aggregation do not correspond with changes in labile organic C fractions in soil amended with 14C-glucose. Soil Biology and Biochemistry., 28: 453-462.
Degens, BP (1998a). Decrease in microbial functional diversity do not result in corresponding changes in decomposition under different moisture conditions. Soil Biology and Biochemistry., 30: 1989-2000.
Degens, BP (1998b). Microbial functional diversity cab be influenced by the addition of simple organic substrates to soil. Soil Biology and Biochemistry., 30: 1981-1988.
Degens, BP (1999). Catabolic response profiles differ between microorganisms grown in soil. Soil Biology and Biochemistry., 31: 475-477.
Degens, BP, Schipper, LA, Sparling, GP & Duncan, LC (2001). Is the microbial community in a soil with reduced catabolic diversity less resistant to stress or disturbance? Soil Biology and Biochemistry., 33: 1143-1153.
Degens, BP, Schipper, LA, Sparling, GP & Vojvodic-Vukovic, M (2000). Decrease in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biology and Biochemistry., 32: 189-196.
Degens, BP & Vojvodic-Vukovic, M (1999). A sampling strategy to assess the effects of land use on microbial functional diversity in soils. Aust. J. Soil. Res., 37: 593-601.
Di Giovanni, GD, Watrud, LS, Seidler, RJ & Widmer, F (1999). Comparison of parental and transgenic alfalfa rhizosphere bacterial communities using Biolog GN metabolic fingerprint and enterobacterial repetitive intergenic consensus sequence-PCR (ERIC--PCR). Microbial Ecology., 37, 129-139.
Dick, RP (1994). Capítulo 7: Soil enzyme activities as indicators of soil quality. En: Doran, JW, Coleman, DC, Bezdicek, DF, Stewart, BA (eds.), Defining Soil Quality for a Sustainable Environment, American Society of Agronomy, Madison, EEUU, pp. 107-124.
Dick, WA, Cheng, L & Wang, P (2000). Soil acid alkaline phosphatase activity as pH adjustment indicators. Soil Biology and Biochemistry., 32: 1915-1919.
Frostegard, A, Tunlid, A, Baath, E (1993). Phospholipid fatty acid, composition, biomass and activity of microbial communities from two soil types experimentally exposed to differentheavy metals. Apply Environmental Microbiology., 59: 3605 - 3617.
Falkowski, P, Scholes, RJ, Boyle, E, Canadell, C, Elser, J, Gruber, N, Hibbard, K, Högberg, P, Linder, S, Mackenzie, FT, Moore, B, Pedersen, T, Rosenthal, Y, Seitzinger, S, Smetacek V & Steffen, W(2000). The global carbon cycle: a test of our knowledge of earth as a system. Science., 290: 291-296.
Germida, JJ, Siciliano, SD & Seib, AM (1998a). Phenotypic plastic of Pseudomonas aureofaciens (lac ZY) introduced into and recovery from field and laboratory microcosm soils. FEMS Microbiology Ecology., 27: 133-139.
Germida, JJ, Siciliano, SD, De Freitas, JR & Seib, AM (1998b). Diversity of root-associated bacteria associated with field-grown canola (Brassica napus L.) and wheat (Tricum aestivum L.). FEMS Microbiology Ecology., 26: 43-50.
Graham, PH, Sadowsky, MJ, Tighe, SW, Thompson, JA, Date, RA, Howieson, JG & Thomas, R (1995). Differences among strains of Bradyrhizobium in fatty acid-methyl ester analysis. Can. J. Microbiol., 41: 1038-1042.
Grierson, PF & Adams, MA (2000). Plant species affect acid phosphatase, ergosterol and microbial P Jarrah (Eucalyptus marginata Donn ex Sm.) forest in south-western Autralia. Soil Biology and Biochemistry., 32: 1817-1827.
Krsek, M & Wellington, EMH (1999). Comparison of different methods for the isolation and purification of total community DNA from soil. Journal of Microbiological Methods., 39: 1-16.
Haldeman, DL, Amy, PS, Ringelberg, D, White, DC, Garen, RE, Ghiorse, WC (1995). Microbiological growth and resuscitation alter community structure after perturbation. FEMS Microbiology Ecology., 17: 27-38.
Harry, M, Gambier, B & Garnier-Sillam, E (2000). Soil conservation for DNA preservation for bacteria molecular studies. Eur. J. Soil Biol., 36: 51-55.
Harry, M, Gambier, B, Bourezgui, Y & Garnier-Sillam, E (1999). Evaluation of purification procedures for DNA extracted from organic rich samples: interference with humic substances. Analusis., 27: 439-442.
Harry, M, Jusseaume, N, Gambier, B & Garnier-Sillam, E (2001). Use of a RAPD markers for the study of microbial community similarity from termite mounds and tropical soils. Soil Biology and Biochemistry., 33: 417-427.
Ibekwe Abasiofiok, M & Kennedy, AC (1998).. Phospholipid fatty acids profiles and carbon utilization patterns for analysis of microbial community structure under field and greenhouse conditions. FEMS Microbiology Ecology., 26: 151-163.
Insam, H (2001). Developments in soil microbiology since the mid 1960. Geoderma., 100: 389-402.
Insam, H & Öhlinger, R (1995). Capítulo 20: Ecophysiological Parameters. En: Schinner, F, Ölhinger, R, Kandeler, E & Margesin, R (eds.), Methods in Soil Biology, Springer Verlag, Berlin, Alemania, pp. 306-309.
Jansa, J, Gryndler, M & Matucha, M (1999). Comparison of the lipid profiles of arbuscular mycorrhizal (AM) fungi and soil saprophytic fungi. Symbiosis., 26: 247-264.
Jarvis, BDW & Tighe, SW (1994). Rapid identification of Rhizobium species based on cellular fatty acid analysis. Plant and Soil., 161: 31-41.
Joergensen, RG (2000). Ergosterol and microbial biomass in the rhizosphere for grassland soils. Soil Biology and Biochemistry., 32: 647-652.
Kandeler, E, Tscherko, D, Bardgett, RD, Hobbs, PJ, Kampichler, C & Jones, TH (1998). The response of soil microorganisms and roots to elevated CO2 and temperature in a terrestrial model ecosystem. Plant and Soil., 202: 251-262.
Kato, M & Wickner, W (2001). Ergosterol is required for the Sec18/ATP-dependent priming step of homotypic vacuole fusion. The EMBO Journal., 20: 4035-4040.
Klavins, M, Dipane, J & Babre, K (2001). Humic substances as catalysts in condensation reactions. Chemosphere., 44: 737-742.
Lechavalier, MP (1977). Lipids in bacterial taxonomy - a taxonomist's view. Critical Review of Microbiology., 5: 109 - 210.
Liu, K (1994). Preparation of fatty acid methyl esters for gas chromatographic analysis of lipids in biological materials. JAOCS., 71: 1179-1187.
Maire, N, Borcard, D, Laczkó, E & Matthey, W (1999). Organic matter cycling in grassland soils of Swiss Jura mountains: biodiversity and strategies of the living communities. Soil Biology and Biochemistry., 31: 1281-1293.
Mc Lean, EO (1982). Soil pH and lime requirement. En : Page AL, Miller RH, Keeney DR (eds). Methods of Soil Analysis, parte 2. Am Soc. Agron., Soil Sci. Soc. Am., Madison, Wisconsin, pp 199 -224.
MIDI (1990). Identification of bacterial by gas chromatography of celular fatty acids, Technical note 101, MIDI, Newark, USA.
Nielsen, P & Petersen, SO (2000). Ester-linked polar lipid fatty acid profiles of soil microbial communities: a comparison of extraction methods and evaluation of interference from humic acids. Soil Biology and Biochemistry., 32: 1241-1249.
Olsson, S & Persson, P (1999). The composition of bacterial populations in soil fractions differing in their degree of adherence to barley roots. Applied Soil Ecology., 12: 205-215.
Olsson, S, Alström, S & Persson, P (1999). Barley rhizobacterial population characterized by fatty acid profiling. Applied Soil Ecology., 12: 197-204.
Ovreas, L, Jensen, S, Daae Frida, L & Torsvik, V (1998). Microbial community changes in a perturbed agricultural soil investigated by molecular and phisiological approaches. Applied and Environmental Microbiology., 64: 2739-2742.
Parkinson, D, Gray, TRG & Williams, ST (1971). Ecology of Soil Microorganisms, Blackwell, Oxford, 116 pp.
Roose-Amsaleg, CL, Garnier-Sillam, E & Harry, M (2001). Extraction and purification of microbial DNA from soil and sediment samples. Applied Soil Ecology., 18, 47-60.
Rosado, AS, Duarte, GF, Seldin, L & Van Elsas, JD (1997). Molecular Microbial ecology: a minireview. Revista de Microbiología., 28: 135-147.
Rundgren, S, Andersson, R, Bringmark, L, Gustafsson, K, Josefsson, M & Torstensson, L (1998). Integrated soil analysis: a swedish research project. Ambio., 27: 2-3.
Ruzicka, S, Edgerton, D, Norman, M & Hill, T (2000). The utility of ergosterol as a bioindicator of fungi in temperate soils. Soil Biology and Biochemistry., 32: 989-1005.
Schinner, F, Öhlinger, R, Kandeler, E & Margesin, R (1996). Methods in Soil Biology. Springer-Vellag, Berlin Heidelberg, Alemania. 426 pp.
Scholes, RJ, Skole, D & Ingram, JS (1995). A global database of soil properties: proposal for implementation. IGBP- DIS Working Paper Nº10.
Schutter, ME & Dick, RP (2000). Comparison of fatty acid methyl ester (FAME) methods for characterizing microbial communities. Soil Science Society of America Journal., 64: 1659-1668.
Siciliano, SD, Theoret, CM, De Freitas, JR, Hucl, PJ & Germida, JJ (1998). Differences in the microbial communities associated with the roots of different cultivars of canola and wheat. Can. J. Microbiol., 44: 844-851.
Siciliano, SD & Germida, JJ (1999). Taxonomic diversity of bacteria associated with the roots of field- grown transgenic Brassica napus cv. Quest, compared to non-transgenic B.napus Excel and B. napa cv. Parkland. FEMS Microbiology Ecology., 29: 263-272.
Sparling, GP & Degens, BP (1999). Soil microbial biodiversity. http://www.landcare.cri.nz/conferences/ manaakiwhenua/papers/index.shtm.
Stenberg, B (1997). Chemical and biological properties of agricultural soil. PhD Thesis/Dissertation University of Uppsala, Sweden. 125 pp.
Stenberg, B (1999). Monitoring soil quality of arable land: microbiological indicators. Acta Agric.Scand., sect B., Soil and Plants Sci., 49: 1-24.
Sundh, I, Börjesson, G & Tunlid, A (2000). Methane oxidation and phospholipid fatty acid composition in a podzolic soil profile. Soil Biology and Biochemistry., 32: 1025-1028.
Tiedje, JM, Asuming-Brempong, S, Nisslein, K, Marsh, TL & Flynn, SJ (1999). Opening the black box of soil microbial diversity. Applied Soil Ecology., 13: 109-122.
Torsvik, VL (1980). Isolation of bacterial DNA from soil. Soil Biology and Biochemistry., 12: 15-21.
Torsvik, VL, Goksoyr, J & Daae, FL (1990). High diversity in DNA soil bacteria. Applied en Environmental Microbiology., 56: 782-787.
Turco, RF, Kennedy, AC, Jawson, MD, (1994). Capítulo 5: Microbial indicators of soil quality. En: Doran, JW, Coleman, DC, Bezdicek, DF & Stewart, BA (eds.), Defining Soil Quality for a Sustainable Environment, American Society of Agronomy, Madison, EEUU, pp. 73-90.
Van Elsas, JD, Duarte, GF, Rosado, AS & Smalla, K (1998). Microbial and molecular biological methods for monitoring microbial inoculants and theirs effects in the soil environment. Journal of Microbiological Methods., 32: 133-154.
Waldrop, MP, Balser, TC & Firestone, MK (2000). Linking microbial community composition to function in a tropical soil. Soil Biology and Biochemistry., 32: 1837-1846.
Widmer, F, Flieábch, A, Laczk¢, E, Schulze-Aurich, J & Zeyer, J (2001). Assessing soil biological characteristics: a comparison of bulk soil community DNA, PLFA and BIOLOG analyses. Soil Biology and Biochemistry., 33: 1029-1036.
Zelles, L (1999). Fatty acids patterns of phospholipids and
lipopolysaccarides in the characterization of microbial communities in soil:
a review. Biol. Fertil. Soils., 29. 111-129.
Zelles, L, Bai, QY, Beek, T & Beese, F (1992). Signature fatty acids in
phospholipids and lipopolysacharides as indicators of microbial biomass and
community structure in agricultural soils. Soil Biology & Biochemistry.,
24: 317-323.
Zogg, GP, Zak, DR, Ringberg, DB, MacDonald, NW, Pregitxer, KS
& White, DC (1997). Compositional and functional shifts in microbial communities
due to soil warming. Soil Science Society of America Journal., 61: 475 - 481.
Sitios WWW de interés
Composición del perfil de fosfolípidos y la tolerancia a metales pesados de las comunidades microbianas del suelo a lo largo de dos gradientes de bosques de coníferas contaminados con metales pesados. [texto en inglés]
Estudio de la dinámica de las comunidades microbianas de los puntos de acopio del estiércol mediante el análisis del perfil de fosfolípidos microbianos. [texto en inglés]
Cambios en los perfiles de fosfolípidos microbianos durante fases de hambruna o desecación de suelos. [resumen en inglés]
Asociación de microorganismos al metabolismo del fenantreno en suelos mediante el análisis de lípidos marcados con 13C. [texto en inglés]
Estudio basado en el patrón de ácidos nucleicos, para evaluar el efecto de la diseminación de la capacidad degradadora plasmidial del 2,4-D sobre la degradación del 2,4-D y la estructura comunitaria bacterial en dos horizontes diferentes del suelo. [texto en inglés]
Estudio basado en el patrón de ácidos nucleicos, para evaluar los efectos de la acidez del suelo sobre la biodegradación de Chlorpyrifos y el aislamiento de bacterias degradadoras de Chlorpyrifos. [texto en inglés]
Estudio sobre si el tipo de suelo es el primer determinante de la composición de las comunidades totales y activas de las bacterias en suelos arables.[texto en inglés]
Libro sobre los Microorganismos como indicadores de la salud del suelo. Informe realizado por el Instituto Nacional de Investigación Ambiental, Dinamarca (en inglés).
Trabajo sobre el flujo de carbono desde las raices a la microbiota del suelo, realizado por la Universidad de California (poster en inglés).
Glosario
Analito - elemento o compuesto que se desea determinar mediante un análisis químico.
Antrópicos - término empleado para denominar aquellas acciones
o productos derivados y/o asociados a la actividad Humana.
Cluster - método estadístico que permite la agrupación
y el ordenamiento de muestras (suelos, en el caso mencionado en el artículo)
basado en los resultados obtenidos para un grupo de variables definidas. En
el caso citado, el patrón de resultados de BIOLOG, PLFA o ADN de cada
suelo estudiado correspondería al grupo de variables utilizadas para
realizar este tratamiento estadístico.
Ecosistema - unidad fundamental de la biósfera que representa un nivel de organización que funciona como un sistema en el que se manifiestan conjuntamente los factores abióticos - físicos y químicos -, así como los factores bióticos (los que provienen de los seres vivos).
Edáfico - (del griego "edáfos", que significa suelo o tierra) término empleado para calificar aquello vinculado con el suelo.
Holística - que pertenece o está asociado con el Holismo. Forma de análisis que indica que las partes no pueden comprenderse separadamente del todo y que los todos son diferentes de la suma de sus partes.
Patrón de restricción - es un conjunto de trozos de ADN, cada uno con diferente tamaño. El patrón de restricción es el producto del ataque de un segmento de ADN con enzimas especiales (llamadas enzimas de restricción) capaces de realizar cortes sobre secuencias específicas de nucleótidos presentes en este ADN. Por lo cual, se espera que de dos ADN idénticos se obtengan patrones de restricción similares.
Transgénicos - término que se emplea para denominar a todas aquellas formas de vida que han sido genéticamente modificadas por el hombre, empleando técnicas de biología molecular.
Xenobiótico - calificativo utilizado para denominar a todos los productos,
físicos y químicos, que son sólo sintetizados por Hombre
y que se han introducido en un ecosistema (por ejemplo: plaguicidas, envases
plásticos, residuos sólidos urbanos, basura radioactiva).