
CELLULAR BIOLOGY |
Train trip into the cell: the Golgi Apparatus as the central station of the intracellular membrane traffic |
[RESUMEN] ABSTRACT
Cellular membranes are crucial for the
cells. The eukaryotic cell is made up of the plasma membrane that isolates it
from the external world, and of intracellular membranes, which form
compartments that give rise to the organelles. This subdivision allows the cell
to adapt to changing external conditions, and to perform specific functions
depending on the tissue. Cell membranes are made up of (glyco)lipids and
(glyco)proteins that once synthesized are sorted and transported to their
specific subcellular destination. Mistakes in this process may compromise cell
survival and, consequently, the whole organism. Hence, the control and
regulation of intracellular transport is a crucial process for the survival of the
cell. The Golgi complex, or Golgi apparatus, is the organelle responsible for
such regulation and control. It can be compared to the Grand Central station on
the intracellular rail trip in which travellers, carriages, wheels, traffic
lights, motors and tracks help us to understand how the intracellular membrane
traffic takes place.
Introduction
The cell is protected from external environment by a surrounding membrane
called plasma membrane. In certain cell types, this membrane presents domains
that are clearly differentiated in function and morphology (cilia, flagella,
microvilli), e.g. polarized cells like neurones and epithelial cells (renal and
intestinal). Other cell types lack these specialized structures e.g.
non-polarized cells like fibroblasts.
The functional
evolution of the eukaryotic cell is parallel to the appearance of new
intracellular compartments. This structural and functional compartmentalization
is determined by the different distribution of the cell membrane components,
lipids and proteins. Their correct distribution in the plasma membrane and in
intracellular membranes allows the cell to fulfil its genetically determined
functions properly. Otherwise serious pathological disorders would take place.
Therefore, the knowledge of the signals that guide lipids and proteins to their
proper destinies (endoplasmic reticulum, Golgi apparatus, lysosomes,
chloroplasts, mitochondria, peroxisomes, plasma membrane) is crucial to
understand the function of the cell. This line of research is included in the
field that studies the intracellular traffic of membranes.
The great routes
of the intracellular traffic of membranes
The intracellular
traffic of membranes directs lipids and proteins to their destination. It has
several pathways (Fig. 1):
1. The secretory, biosynthetic
or exocytic pathway. This
intracellular route is followed by the molecules synthesized in the endoplasmic
reticulum (ER) that are transported to (a) other subcellular compartments (such
as the Golgi apparatus, the lysosomes, the cloroplats, etc), (b) the plasma
membrane, and (c) the extracellular medium. We distinguish two types of
secretion:
1.1. Constitutive secretion. As soon as lipids and proteins are
synthesized, they are continuously transported and secreted to their final
destination. This secretion happens in all cells. An example for a protein that
follows is contitutively secreted is given in a recent review by Ureña &
Arribas (2000)[1].
1.2. Regulated secretion. It takes place only in response to specific
signals e.g. certain ions or as a consequence of the hormone-receptor
interaction. Once the products susceptible to regulated secretion have been
synthesized, they are stored in spherical membrane structures called vesicles
or granules, depending on their size. This secretion is only reported in
certain cell types such as endocrine and exocrine cells, macrophages, some
leukocytes and neurons.
2. The endocytic pathway. This is the route by which soluble and
membrane components enter into the cell. This includes:
2.1. The receptor-mediated endocytosis.
The molecules that enter into the cell first bind to either a cell surface
receptor or to a receptor stored in intracellular compartments that rapidly
migrate to the plasma membrane in response to specific signals. For instance,
the glucose receptor GLUT4 is stored in vesicles just localized bellow the
plasma membrane. Increase in the plasma concentration of glucose leads to the
secretion of insulin, which binds to cell surface receptors. This binding
induces the vesicles to fuse with the plasma membrane where the receptors are
incorporated. Free glucose is then taken up by these receptors and both
re-enter the cell, glucose is then released and GLUT4 receptors can be used in
a new cycle. The uncoupling between the this signalling and trafficking
processes results in insulin-independent diabetes mellitus.
2.2. Endocytosis by pinocytosis. This allows the entry of macromolecules and fluids, which renews the plasma membrane every 30-60 min.
2.3. Endocytosis by caveolae. This mechanism uses vesicles that carry mainly caveolin and capture small hydrophobic molecules such as cholesterol and folic acid. Caveolae are involved in intracellular signal transduction.
2.4. Phagocytosis is a specialized form of endocytosis that incorporates large particles such as viruses, bacteria, intracellular parasites and inert complexes. It takes place only in certain cell types such as macrophages and neutrophils.
3. The recycling pathway. Certain membrane components enter the cell but once the load is released, they are returned to the plasma membrane so that they can be used again. This is what happens with most membrane receptors. In fact, it is a combination of the endocytic (internalising) and the secretory (return to the cell surface) pathways.
Regarding the amount of intracellular membrane, secretion and endocytosis are highly equilibrated. Any alteration would compromise cell survival. However, we will not focus on the mechanisms that are involved or that regulate these pathways, since they are beyond the scope of this text.
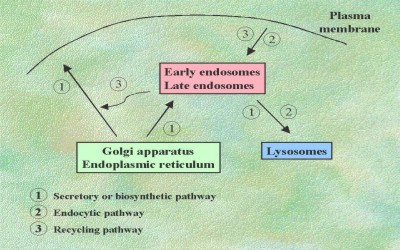
Fig.1. Intracellular pathways of membrane trafficking.
Train trip into
the cellTrain trip into the cell
The organelles
and molecular components involved in membrane traffic can be compared to a
train journey and itemized as follows: passengers would stand for lipids and
proteins, carriages for transport vesicles and stations for organelles
(endoplasmic reticulum, lysosomes, endosomes, plasma membrane). We will focus
on the intracellular Grand Central Station: the Golgi apparatus.
The endoplasmic reticulum as the hangar,
factory and assembly line of intracellular trains
Lipids and
proteins are synthesized in the ER, which consists of a continuous net of
cisternae covered by ribosomes and extends through all the cytoplasm. Protein
synthesis occurs on ribosomes, where the genetic message encoded in mRNA is
translated. mRNA results from the transcription of DNA in the nucleus and is
then transported to the cytoplasm. It encodes for cellular proteins, which may
remain in the cytoplasm or else be transferred to the ER. Among the latter,
some are free or soluble inside the ER cisternae (lumen) and some are
membrane-bound. They could be referred to as standing passengers (soluble or
luminal proteins) and seated passengers (membrane proteins). They comprise on
the one hand proteins of the ER, which have to be retained, and on the other
hand proteins that must end up in other compartments to function and proteins
that are to be secreted to the outside of the cell.
All proteins have
a three-dimensional structure that is achieved by a continuous folding. Only
properly folded proteins can be transported. Spontaneous twisting often brings
about errors that may inactivate the protein. To prevent this from happening, a
set of proteins of the ER lumen called chaperones help their mates in the
contortion exercises. They contribute to the slow and ordered folding of
proteins and to the packaging of their different subunits (Ellgaard et al,
1999). However, the most essential item in a journey is the ticket. What is the
ticket? Which passengers are allowed to travel and which not? Unlike real
passengers, certain molecular passengers can travel free! However, others must
pay. What does this mean? According to the bulk-flow theory of exit (Wieland et
al, 1987), the departure of soluble and membrane-bound proteins (both termed as
cargo) from the ER does not need specific signals. However, recent data suggest
that certain proteins present a signal sequence at the carboxyl terminal end that
consists of two phenylalanines or, at least, two acidic amino acids (Nishimura
& Balch, 1997). The signal sequences that determine whether the ER proteins
stay there are better characterized: di-lysine and di-arginine are required at
the carboxyl terminal end for type I and type II membrane proteins,
respectively (Nilsson et al, 1989; Jackson et al, 1990; Schutze et. al, 1994),
and lysine-asparagine-glutamic acid-leucine (KDEL) at the carboxyl terminal end
for soluble proteins (Munro & Pelham, 1987). In principle, all the proteins
that lack these ER retention signals will leave the ER towards the Golgi
apparatus. However, the ER and the Golgi are not physically connected and so
membrane intermediaries or transport vesicles are needed to make this intracellular
journey.
Transport vesicles or the formation and
assembly of intracellular wagons
Both the soluble
and membrane-bound proteins that follow the secretory pathway leave the ER
towards the Golgi apparatus, where their molecular structure is completed
(glycosylation, phosphorylation) and they are packaged and sorted to their
appropriate final destination. Export from the ER is extensively studied and
there are more data on the molecular machinery involved than on its regulation.
The ER is bigger than the Golgi apparatus, which means that the ER proteins are
diluted and must concentrate throughout their journey to the Golgi. This is
achieved in parallel to the formation of transport vesicles, which takes place
in specific ribosome-free areas of the ER called exiting sides. Thus not all
the ER is able to accumulate cargo and form transport vesicles. The wagons
(vesicles) can only be found at platforms (exiting sides).
Two relevant
facts are associated with transport: (1) the loading of the cargo and (2) the
deformation of the membrane at the exit sites and the subsequent separation
that lead to the formation of the vesicle. Passengers (proteins and lipids) are
scattered around the station and when the time of departure of the train
(vesicles) approaches, they gather on the platform (exiting sites). The
auto-assembly of several coats of multiproteic complexes deforms the donor
membrane to form COP (coat proteins)-coated vesicles. There are COPI- and
COPII-coated vesicles (Kreis and Pepperkok, 1994). These vesicles are generated
in tandem (Nickel et al., 1998), i.e. COPII vesicles are originated in the ER
first, then they fuse to each other giving rise to the vesicular-tubular
transport complex (VTC) from where COPI vesicles are formed. These VTCs are
considered a labile pleomorphic organelle that is also known like endoplasmic
reticulum-Golgi intermediate compartment (ERGIC; Hauri et al, 2000) (Fig. 2).
The VTC or ERGIC is a mobile membrane structure that carries and concentrate
secretory components (Martínez-Menárguez et al, 1999). For further details
about the molecular machinery required for the formation of COP-coated
vesicles, see specialized reviews (Rothman & Wieland, 1996; Schekman and
Orci, 1996; Wieland & Harter, 1999; Springer et al, 1999). There is another
coat called clathrin that operates in the intracellular traffic. It is the
association of three heavy and three light chains that form the so-called
tryskelion units and are linked as polyhedrons like the net in basketball. They
were first associated with receptor-mediated endocytosis but they have also
been reported in the transport of proteins from the Golgi apparatus to
lysosomes (Le Borgue & Hoflack, 1998).
The formation of
vesicles is regulated to a great extent by other molecules, most of which are
also directly involved in intracellular signaling processes (Stow, 1995; De
Camilli et al, 1996). This allows the cell to regulate its intracellular
traffic according to specific external signals, just as given events (sports,
political, musical, etc) and times of the year (height of the season, off
season) determine the travel demand. In order to meet such demand, the number
of wagons, their size and the frequency of trains have to be carefully
regulated so that they neither stop the traffic nor waste membranes or energy
since the formation of vesicles requires ATP.
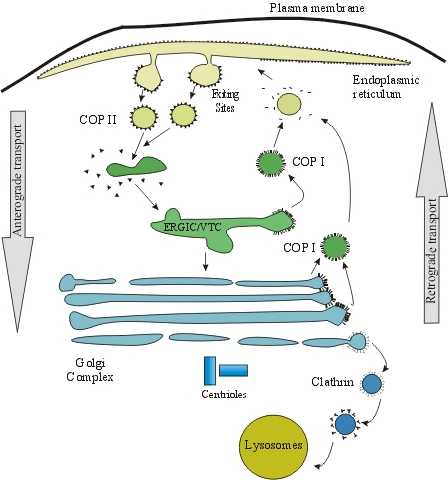
Fig. 2. Diagram of the intracellular transport in the
endoplasmic reticulum and the Golgi apparatus. The latter is surrounded by the
ER and is organized around the centrioles. Between the Golgi and the ER,
several tubular and vesicular structures form a pleomorphic and dynamic
compartment termed as ERGIC (endoplasmic reticulum-Golgi intermediate
compartment) or VTC (vesicular-tubular transport complex).
The Golgi apparatus: the Grand central station of intracellular traffic in eukaryotic cells
In most
non-polarized animal cells, the Golgi apparatus is an active and a single-copy
organelle. It consists of a set of stacks made up of highly flat cisternae with
dilated rims. These stacks are interconnected by means of tubules and vesicles
(Rambourg y Clermont, 1990). The Golgi apparatus is responsible for most the
post-translational modifications of proteins (Driouch & Staehelin, 1997;
Farquhar & Palade, 1998). The most frequent is glycosylation that gives
rise to glycolipids and glycoproteins, and it consists in extensive
modifications of the carbohydrate groups (glycans or saccharides) that have
been previously added en block to the proteins synthesized in the ER.
Phosphorylation is also another post-translational modification that occurs in
the Golgi apparatus, which is essential for the sorting of luminal proteins to
lysosomes, as well as the sulfatation of proteoglycans and some proteolytic
reactions that are crucial for certain hormones.
In animal cells,
the Golgi apparatus is localized close to the nucleus and around the centrosome
(the cytoplasmic organelle where microtubules are formed) (Fig. 3A). The ER
surrounds the Golgi apparatus (Fig. 2) (Note: in the Cell and Molecular Biology,
and Biochemistry textbooks, the Golgi is located between the ER and the plasma
membrane to simplify the secretory pathway, which does not reflect the real
intracellular arrangement in non-polarized cells, frequently leading to errors
of interpretation).
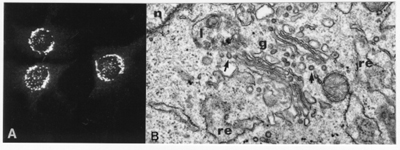
Fig. 3. (A) The Golgi apparatus of
mammalian cells cultured with antibodies against one of its resident protein
(mannosidase II) as visualized under the fluorescence microscope. The Golgi
shows a reticular morphology that extends around the nucleus. (B) The Golgi
apparatus (g) seen under the transmission electron microscope. This organelle
consists of a stack of flat cisternae and is surrounded by ribosome-attached
cisternae from the endoplasmic reticulum (re). The arrows show COPI-coated
vesicles.(n, nucleus; l, lysosome).
In both animal
and plant cells, a stack of the Golgi consists of flat cisternae (central zone)
with an entry (cis) and an export area (trans) (Fig. 3A). Each area is linked
to a net of tubular and vesicular structures: one of entry (cis-Golgi network,
CGN) and one of exit (trans-Golgi network, TGN) (Rambourg y Clermont, 1990)
(Fig. 2). The morphologic polarity of the Golgi apparatus results in functional
polarity and vectorial traffic. During this journey, the lipids and proteins passing
through the Golgi undergo several sequential modifications determined by the
molecular composition of the enzymes unevenly distributed in the cisternae
(Roth, 1997; Varki, 1998). In contrast to the well-characterized signal
sequences of the ER, the sequences added by glycosyltransferases or the
structural requirements that allow their retention in specific cisternae of the
Golgi are unclear.
Assembly and maintenance of the Golgi apparatus in eukaryotic cells. Vesicular and cisternal-maturation transport models.
The Golgi apparatus is also composed by membranes.
Therefore, in spite of the intense entry and export traffic, its size and shape
remain constant. This requires a finely regulated equilibrium of the input and
output flows to prevent the hypertrophy and the atrophy of the Golgi, which
would compromise the cell survival. Thus the integrity of the Golgi apparatus
results from the equilibrium between the incoming (anterograde) and the
outgoing (retrograde) traffic (Fig. 2). The former is the membrane flow that
enters and is then destined for the plasma membrane. The latter refers to the
flow that arrives at and originates in the Golgi or passes through it and is
then directed to the ER. The retrograde route is used by membrane and soluble
proteins that have left the ER for the Golgi by mistake and then should
returned to the ER. To this end, they need specific receptors that recognize
the motifs for ER retention (the receptors of the K(H)DEL sequence mentioned
above) (Lewis & Pelham, 1990). It is also the pathway used by certain
toxins (e.g. the cholera and Shiga toxins) to reach the ER and exert their
toxic effect. In this case, the Golgi apparatus is an unavoidable checkpoint
before reaching the ER (Sandvig et al, 1992).
However, what determines and regulates the specificity and
consistency of both flows? Or, in other words, which are the traffic lights and
the points that govern the arrival and departure of intracellular trains? The
specificity of recognition and of membrane fusion is due to the molecular
interaction between a series of multiproteic complexes, as suggested by the
SNARE hypothesis of vesicular transport (Rothman and Warren, 1994). According
to this model, there is a molecular interaction between a group of proteins
such as the NSF (NEM-sensitive fusion protein), the SNAPs (soluble NSF-binding
proteins) and the SNAREs (receptors of SNAPs). The latter are present at both
the donor (vSNARE) and the acceptor (tSNARE) membranes, such as a transport
vesicle and a Golgi cisternae or plasma membrane, respectively. The interplay
between vSNARE and tSNARE should be unique and should ensure the specifity of
the fusion. Several SNAREs have been identified in certain compartments. They
could be regarded as the points that switch each train to its corresponding
platform. However, they are not totally specific and so require the help of a
family of proteins termed Rabs (Novick and Zerial, 1997). These proteins attach
to membranes after activation by binding to GTP. Each compartment has a
specific Rab protein, which represents the signal that avoids train collisions
during the switching of rails and errors in the location of trains at
platforms. There are other minor traffic regulators but they are beyond the
scope of this text. Whatever the direction, transport is mediated by vesicles
(Rothman, 1994) (Fig. 4A). The size of these structures is 60-80 nm and the
SNARE and Rab proteins determine their specificity and consistency. The
vesicular model is a good candidate for the transport of most lipids and
proteins, since the sizes agree. Moreover, the volume/area ratio is high, which
increases the carrying capacity. However, the cell also secretes large
molecules and proteins e.g. the scales of certain algae, the procollagen
synthesized by fibroblasts and the multiproteic complexes between apoprotein E
and albumin in the hepatocyte, none of which fits in vesicles. In these cases,
an alternative model has been suggested: the cisternal-maturation (Fig. 4B)
(Mironov et al, 1997; Glick and Malhotra, 1998). According to this hypothesis,
a cisterna is formed in the cis site of the Golgi apparatus by continuous
fusion of VTCs formed in the exiting sites of the ER (Fig. 4B). This results in
the cis-most cisterna of the Golgi. Posterior transport to the plasma membrane
is favored by the progressive shift of cisternae to the trans site of the
Golgi. Transported proteins in such a way would never be removed from cisternae
and Golgi-resident proteins (glycosyltransferases) would be transported back to
adjacent cisternae by vesicles. Finally, in the trans-most site of the Golgi
(TGN), the mature cisterna would either be transported as such or split giving
rise to tubules in order to merge with the plasma membrane.
Mitosis
is frequently used as a model for the study of the molecular mechanisms of
assembly and disassembly of the Golgi apparatus (Warren & Malhotra, 1998).
This is the process of division that results in the production of daughter
cells genetically identical to the parent cell from which they arose and with the
same organelles. During mitosis, the Golgi apparatus is extensively fragmented
and homogenously distributed through the cytoplasm so that it is equitably
distributed to each daughter cell. At the end of the division, these fragments
are re-assembled to achieve the classical morphology of the Golgi (Lowe et al,
1998). However, this is exclusive to mammalian cells. In yeast, another major
model for the study of the molecular mechanisms involved in intracellular
traffic (Duden & Schekman, 1997), Drosophila (Stanley et al., 1997), and
plant cells (Driouich & Staehelin, 1997) the Golgi virtually remains
unaltered during mitosis.
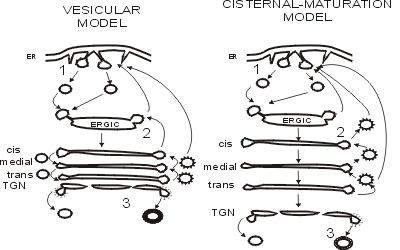
Fig. 4. General diagram of two models of intracellular
transport: the vesicular (A) and the cisternal-maturation (B) models. See text
for further details. Partially adapted from Glick and Malhotra (1998).
High- and low-speed routes for intracellular transport: the cytoskeleton and its association with the Golgi apparatus
In sum,
intracellular traffic is mainly mediated by vesicles, which requires the
sorting of the cargo in the specific membrane domains of each compartment,
followed by transport and subsequent fusion of the vesicle and the membrane of
the receptor compartment.. We have also described the molecular mechanisms that
regulate these processes (COPI and COPII coats; SNAREs, rabs). In addition,
another set of proteins establish and maintain the shape of compartments,
retain the organelles at specific sites of the cell and favour the fusion and
movement of vesicles so that they end up in the appropriate compartments. We
are going to talk now about motors, wheels and rails.
All
cells present a cytoskeleton that is involved in the subcellular organization
of the cell, that maintains the shape of the cell, and it consists of microtubules
(Fig. 5A), intermediate filaments and actin microfilaments (Fig. 5B).
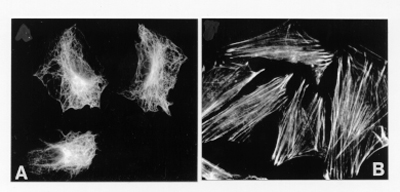
Fig.
5. The cytoskeleton in eukaryotic cells. (A) The microtubular net as visualized
using anti-b-tubulin antibodies. (B) The actin cytoskeleton (stress fibres) as
visualized using the phalloidin toxin.
Microtubules
are linear structures composed of tubulin units that point to the periphery of
the cell from the centriole. As has been pointed out, the Golgi apparatus is
located around centrioles (Fig. 5A) centralizing the membrane flow originated
in the ER (Fig. 2) (Cole and Lippincott-Schwartz, 1995). The Golgi apparatus is
connected to cytoskeleton (Kreis et al, 1997). The factors that affect the
structure of microtubules also alter the integrity and localization of the
Golgi apparatus (Fig. 5A) by splitting it into small pieces or ministacks that
are dispersed throughout the cytosplasm (Fig. 5B). Both the Golgi and the
transport vesicles interact with microtubules through a family of proteins
called dyneins and kinesins, which are able to transform ATP into movement.
They are thus referred to as motor proteins (Allan, 1996). In non-polarized
cells, dynein allows movement towards the centrosome, whereas kinesin do it to
the plasma membrane (Lane & Allan, 1998). It is just as the wheels
(vesicles) move on the rails (microtubules). Since this is a direct and rapid
means of transport, microtubules represent the intracellular high-speed routes.
However, the Golgi apparatus remains attached to microtubules when the motor
function is inactivated. This shows that there must be a specific set of
proteins to ensure a continuous attachment between the Golgi and the
microtubules (Infante et al, 1999). These proteins may behave as the brakes of
organelles and, in this case, of the Golgi apparatus.
On the
other hand, the Golgi also interacts with the actin microfilaments (Valderrama
et al, 1998, 2000). Microfilaments are also composed of polymerized actin units
and their diameter and length are lower than those of microtubules and they
have numerous ramifications. They thus form a dense, flexible, dynamic
cytoplasmatic network that is responsible for the motion of cells. The breaking
of actin microfilaments brings on the compactness of the Golgi apparatus (Fig.
5C).
Fig. 6. The subcellular localization and morphology of
the Golgi apparatus (A) depend on the cytoskeleton. The disruption of
microtubules by nocodazole (noc) leads to the fragmentation of the Golgi and
its spreading throughout the cytoplasm (B), whereas the disruption of the actin
microfilaments by cytochalasin D (cyD) leads to its compactation (C).
Microfilaments
are also associated with another type of motor proteins termed myosins
(Sellers, 1999). The rate of transport through microfilaments (low-speed route)
is lower than through microtubules. However, the reticular organization allows
access to every area of the cell, reminiscent of suburban trains, which cover
short distances and have frequent stops.
Finally,
the Golgi also contains spectrin and ankyrin isoforms (Holleran and Holzbaur,
1998), which are thought to behave as scaffolds. The cytoskeleton of the
erythrocytesis based upon spectrin, which accounts for the flexibility of these
cells in passing the narrow capillaries of the blood microflow. However,
although the role of such proteins in the Golgi apparatus is unknown, they
could be involved in the flat shape of cisternae rather than in intracellular
transport.
APENDIX
Is the Golgi apparatus a dependent or independent organelle with respect to the ER?
Recent data on the Golgi apparatus have given rise to a strong controversy. The Golgi apparatus was understood to be an organelle that was tightly associated with the ER, with which it maintained a fine dynamic membrane balance. However, two recent articles suggest that this is not the case, and that the Golgi is an independent organelle. If the biogenesis and physiology of the Golgi apparatus depend on the ER, one might assume that a functional Golgi could be re-built from the ER. Using microsurgery techniques, Pelletier et al. (2000) obtained subcellular fragments that contained solely ER membranes. They observed that such fragments were able to synthesize and transport cargo out the ER. However, this cargo was not secreted but retained in the ER exiting sites since no Golgi membranes were present in these subcellular fragments. To assess whether the ER was essential to the formation of the Golgi apparatus, in another study Seemann et al. (2000) treated cells with a drug (brefeldin A) that reversibly induces the merging of Golgi and ER membranes. When brefeldin A was withdrawal in cells that expressed a negative mutant of Sar1 (a crucial protein for the formation of COPII-coated vesicles from the ER), they observed the appearance of a Golgi-like structure that contained known Golgi matrix proteins but was devoid of the Golgi glycosyltranferases. The latter were still retained in the exiting sites of the ER as a consequence of the inhibitory effect of the Sar1 mutant protein. Taken together, these experiments indicate that the ER is neither necessary nor sufficient for the formation and function of the Golgi apparatus. Hence, they indicate that this organelle is independent of the ER (Kumplerman, 2000). Immediately afterwards, new Golgi transport models appeared (Pelham and Rothman, 2000; Stephens and Pepperkok, 2001).
Camilo Golgi discovered the Golgi apparatus a little over 100 years ago (Golgi, 1898). Over the last 20 years the field of intracellular traffic has seen tremendous advances towards the identification of the relevant molecular machineries. Proteins involved in budding, fission, fusion, and sorting have been discovered, and, in some cases, a picture of how such proteins are assembled and work has been glimpsed. In contrast, perhaps surprisingly, a satisfactory understanding of how transport occurs in vivo at the organelle level has not been achieved. The novel combination of GFP (green fluorescent protein) technology and advanced higher resolution quantitative video microscopy with correlative light-electron microscopy will doubtless provide new and relevant information about the transport into, through and from the Golgi apparatus (Lippincott-Schwartz et al., 1998; Polishchuk et al., 2000; Keller et al., 2001). We trust we will not have to wait another century for the complete understanding of this complex and dynamic organelle.
Acknowledgements
I thank
the present and past members of my laboratory, Inmaculada Ayala, Teresa Babià,
Juan Manuel Durán, Ana Luna, Olga Matas and Ferran Valderrama, for their
contributions and stimulating discussions, and in particular Jesús Avila,
Carlos Dotti, Gareth Griffiths, Jaime Renau-Piqueras and Vivek Malhotra for
their continuous support and collaboration, and Robin Rycroft for linguistic
assistance. The work in the authors laboratory was supported by CICYT and
CIRIT grants and/or fellowships from the Spanish Ministry of Education and
Science/Culture, the Generalitat de Calatunya, and the University of Barcelona.
This article is dedicated to José Cortina for letting me do all what I wanted
in the laboratory of Sciences when I was a student at the High School and to my
family for their invaluable patience and love.
Pointers of Interest
Citoskeleton
1. TM Terrys Laboratory (Universidad de Connecticut, Storrs)
http://cmgm.stanford.edu/theriot/movies.htm
2. Joel Swanson (Univ. Michigan, Ann Harbor, USA)
http://www.umich.edu/~jswanlab/Movies/movies.html
3. Phagocytosis
http://www.umich.edu/~jswanlab/Images/images.html.
4. Microtubule dynamics during mitosis, motility and membrane
traffiqking (Salmon Lab Movies)
http://www.unc.edu/depts/salmlab/salmonlabmovies.html
Motors
1. Molecular motors (Molecular Motors Group at the University of York, UK)
http://motility.york.ac.uk:85/
2. Kinesine Superfamily (Laboratorio de N.
Hirokawa, Tokyo, Japón)
http://cb.m.u-tokyo.ac.jp/
3. Ron Vales molecular motors lab
http://cmp.ucsf.edu/valelab/
4. Kinesine Movements along a microtubule
http://math.lbl.gov/~hwang/animation/walk9.mpeg
http://www.bio.brandeis.edu/~gelles/kamppnp/index.html
5. Myosine
http://www.mrc-lmb.cam.ac.uk/myosin/myosin.html
6. Myosine II during cell division and migration
http://www.stc.cmu.edu/CLMIBhp/Imggallpg/
7. Force generated by myosine II on an actine microfilament
http://cmgm.stanford.edu/~wshih/gif.html
8. Vesicular transport along skid giant axon
microtubules and microfilaments (Laboratory de M. Langford)
http://www.dartmouth.edu/~langford/
9. Microtubules and microfilaments cellular
motility, axonal growth, and melanophores (Laboratorio de G. Borisy)
http://borisy.bocklabs.wisc.edu/pages/movies.html
10. Complete list of cytoeskeleton labs
http://vl.bwh.harvard.edu/labs.shtml#cytoskeleton
El complejo de Golgi y tráfico intracelular
1. Golgi complex in Plants
http://cs3.brookes.ac.uk/schools/bms/research/molcell/hawes/gfp/gfp.html
2. Images from the Viki Allans lab web page
http://www.biomed2.man.ac.uk/allan/
3. Images from the Jennifer Lippincott-Schwartzs
lab
http://dir.nichd.nih.gov/CBMB/uobhome.htm
http://dir.nichd.nih.gov/cbmb/pb1labob.html
4. Images from the Graham Warrens lab
http://info.med.yale.edu/cellbio/Warren.html
5. Derek Toomres lab
http://www.livingroomcell.com/
6. Rainer Pepperkoks lab
http://www.embl-heidelberg.de/ExternalInfo/pepperko/index.html
http://www.rpi.edu/~plourj/cellbiol/er-golgi.htm
7. Bruno Gouds lab
http://www.curie.fr/sr/cdrom/equipes/goude.htm
5. Kai Simons lab
http://www.mpi-cbg.de/content.php3?lang=en&aktID=simons
http://www.embl-heidelberg.de/ExtrenalInfo/simons/movies.html
6. Felix Wielands lab
http://www.uni-heidelberg.de/zentral/bzh/wieland.html
7. Golgi Apparatus 3D Organization
http://bio3d.colorado.edu/
8. Clatrine mediated endocytosis
http://www.hms.harvard.edu/news/clathrin/index.html
9. Thomas Kreiss lab
http://www.unige.ch/keris-lab/FP/FP_Video.html
Miscellaneous
1. General web page for Cellular biologists (P. Lafont)
References
Allan, V.J. (1996) Role of motor proteins in organazing the endoplasmic reticulum and Golgi apparatus. Sem. Cell Dev. Biol. 7: 335-342.
Allan, V.J. & Schroer, T.A. (1999) Membrane motors. Curr. Opin. Cell Biol. 11: 476-482.
Cole, N.B. & Lippincott-Schwartz, J. (1995) organization of organelles and membrane traffic by microtubules. Curr. Opin. Cell Biol. 7: 55-64.
De Camilli, P., Emr, S.D., McPherson, P.S. & Novick, P (1996) Phosphoinositides as regulators in membrane traffic. Science, 271:1533-1539.
Driouich, A. & Staehelin, L.A. (1997) The plant Golgi apparatus: structural organization and functional properties. In: The Golgi apparatus. Eds. Berger, E.G. & Roth, J. Birkhäuser Verlag, Basel, pp. 275-302.
Duden, R. & Schekman, R. (1997) Insights into Golgi function through mutants in yeast and animal cells. In: The Golgi apparatus. Eds. Berger, E.G. & Roth, J. Birkhäuser Verlag, Basel, pp 219-246.
Ellgaard, L., Molinari, M & Helenius, A (1999) Setting the standards: quality control in the secretory pathway. Science, 286: 1882-1888.
Farquhar, M.G. & Palade, G.E. (1998) The Golgi apparatus: 100 years of progress and controversy. Trends Cell Biol., 8:2-10.
Glick, B.S. & Malhotra, V. (1998) The curious status of the Golgi apparatus. Cell, 95: 883-889.
Golgi, C. (1898) Sur la structure des cellules nerveuses. Arch. Ital. Biol. 30:60-71 (Original publicado en Boll. Soc. med.-chir. di Pavia, 1989).
Hauri, H.-P., Kappeler, F., Andersson, H. & Appenzeller, C. (2000) ERGIC-53 and traffic in the secretory pathway. J. Cell Sci., 113: 587-596.
Holleran, E.A. & Holzbaur, E.L.F. (1998) Speculating about spectrin: new insights into Golgi-associated cytoskeleton. Trends Cell Biol. 8: 21-25.
Infante, C., Ramos Morales, F., Fedriani, C., Bornens, M. & Ríos, RM (1999) GMAP 210, a cis-Golgi network-associated protein, is a minus microtubule-binding protein. J. Cell Biol., 145:83-98.
Jackson, M.R., Nilsson, T. & Peterson, P.A. (1990) Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J., 9:3153-3162.
Keller, P., Toomre, D., Díaz, E., White, J. & Simons, K (2001) Multicolor imaging of post-Golgi sorting and trafficking in live cells. Nat. Cell Biol. 3: 140-150.
Kreis, T.E. & Pepperkok, R. (1994) Coat proteins in intracellular membrane transport. Curr. Opin. Cell Biol., 6: 533-537.
Kreis, T.E., Goodson, H.V., Pérez, F. & Rönnholm, R. (1997) Golgi apparatus-cytoskeleton interactions. In: The Golgi apparatus. Eds. Berger, E.G. & Roth, J. Birkhäuser Verlag, Basel, pp179-194.
Kreis, T.E., Lowe, M. & Pepperkok, R. (1995) COPs regulating membrane traffic. Ann. Rev. Cell Develop. Biol. 11:677-706.
Klumperman, J. (2000) The growing Golgi: in search of its independence. Nat. Cell Biol. 2: E217-219.
Lane, J. & Allan, V.J. (1998) Microtubule-based membrane movement. Biochim. Biophys. Acta 1376: 27-55.
Le Borgue, R. & Hoflack, B. (1998). Mechanisms of protein sorting and coat assembly: insights from the clathrin-coated vesicle pathway. Curr. Opin. Cell Biol., 10: 499-503.
Lewis, M.J. & Pelham, H.R.B. (1990) The human homologue of the yeast HDEL receptor. Nature, 348: 162-163.
Lippincott-Schwartz, J., Cole, N., & Presley, J. (1998) Unraveling Golgi membrane traffic with green fluorescent protein chimeras. Trends in Cell Biol. 8: 16-21.
Lowe, M, Nakamura, N. & Warren, G. (1998) Golgi division and membrane traffic. Trends Cell Biol., 8: 40-45.
Martínez-Menárguez, J.A., Geuze, H., Slot J.W. & Kumplerman, J. (1999) Vesicular tubular clusters between the ER and Golgi mediate concentration of soluble secretory proteins by exclusion from COPI-coated vesicles. Cell, 98:81-90.
Mironov, A.A., Weidman, P. & Luini, A. (1997) Variations on the intracellular transport theme: maturing cisternae and traficking tubule. J. Cell Biol., 138: 481-484.
Munro, S. & Pelham, H.R. (1987) A C-terminal signal prevents secretion of lumenal ER proteins. Cell, 48:899-907.
Nickel, W., Brügger, B. & Wieland, F.T. (1998) Protein and lipid sorting between the endoplasmic reticulum and the Golgi apparatus. Sem. Cell Dev. Biol. 9:493-501.
Nilsson, T., Jackson, M., & Peterson, P.A. (1989) Short cytoplasmic sequences serve as retention signals for transmembrane proteins in the endoplasmic reticulum. Cell, 58: 707-718.
Nishimura, N. & Balch W.E. (1997) A di-acid signal required for selective export from the endoplasmic reticulum. Science, 277:556-558.
Novick, P. & Zerial, M. (1997) The diversity of Rab proteins and vesicle transport. Curr. Opin. Cell Biol., 9:496-504.
Pelham, H. & Rothman, J.E. (2000) The debate about transport in the Golgi two sides of the same coin? Cell 102:713-719.
Pelletier, L. Jokitalo, E. & Warren, G. (2000) The effect of Golgi depletion on exocytic transport. Nat. Cell Biol. 2:840-845.
Polishchuk, R.S., Polishchuk, E.V., Marra, P., Alberti, S., Buccione, R., Luini, A. & Mironov, A.A. (2000) Correlative light-electron microscopy reveals the tubular-saccular ultrastructure of carriers operating between Golgi apparatus and plasma membrane. J. Cell Biol. 148: 45-58.
Rambourg, A. & Clermont, Y. (1997) Three-dimensional structure of the Golgi apparatus in mammalian cells. In: The Golgi apparatus. Eds. Berger, E.G. & Roth, J. Birkhäuser Verlag, Basel.
Roth, J (1997) Topology of glycosylation in the Golgi apparatus. In: The Golgi apparatus. Eds. Berger, E.G. & Roth, J. Birkhäuser Verlag, Basel, pp 131-162.
Rothman, J.E. (1994) Mechanisms of intracellular protein transport. Nature, 372:55-63.
Rothman, J.E. & Warren, G. (1994) Implications of the SNARE hypothesis for intracellular membrane topology and dynamics. Curr. Biol., 4:220-233.
Rothman, J.E. & Wieland, FT. (1996) Protein sorting by transport vesicle. Science 272: 227-234.
Sandvig, K., Garred, O., Prydz, K., Koslov, J.V., Hansen, S.H. & van deurs, B. (1992) Retrograde transport of endocytosed Shiga toxin to the endoplasmic reticulum. Nature, 358:510-512.
Schekman, R. & Orci, L (1996) Coat proteins and vesicle budding. Science, 271:1526-1533.
Schutze, M.P., Peterson, P.A. & Jackson, M.R. (1994) An N-terminal double-arginine motif mantains type II membrane proteins in the endoplasmic reticulum. EMBO J., 14:1329-1339.
Seemann, J., Jokitalo, E., Pypaert, M. & Warren, G (2000) Matrix proteins can generate the higher order architecture of the Golgi apparatus. Nature 407: 1022-1026.
Sellers, J.R. (1999) Myosins (2nd. Edition). Oxford University Press, Oxford, pp. 1-237.
Springer, S., Spang, A. & Schekman, R. (1999) A primer on vesicle budding. Cell, 97: 145-148.
Stanley, H., Bota, J., Tokuyasu, K & Malhotra, V (1997) The mechanism of Golgi seggregation is cell type specific. Proc. Natl. Acad. Sci. USA 94: 14467-14470.
Stow, J.L. (1995) Regulation of vesicular transport by GTP-binding proteins. Curr. Opin. Nephrol. Hypertens., 4:421-425.
Stephens, D.J. & Pepperkok, R. (2001) Illuminating the secretory pathway: when do we need vesicles? J. Cell Sci. 114:1053-1059.
Ureña, J.M.
& Arribas J. (2000). El transporte intracelular de proteinas. El caso del
TGF-a. http://www.ciencia.cl/CienciaAlDia/volumen3/numero1/articulos/articulo2.html).
Valderrama F., Babià, T., Ayala, I., Kok, J.W., Renau-Piqueras, J. & Egea, G. (1998) Actin microfilaments are essential for the cytological positioning and morphology of the Golgi complex. Eur. J. Cell Biol., 76: 9-17.
Valderrama, F., Luna, A., Babià, T., Martínez-Menárguez, J.A., Ballesta, J., Barth, H., Chaponnier, C., Renau-Piqueras, J., & Egea, G. (2000) The Golgi-associated COPI-coated buds and vesicles contain b/g-actin. Proc. Natl. Acad. Sci. USA 97: 1560-1565.
Varki, A. (1998) Factors controlling the glycosylation potential of the Golgi apparatus. Trends Cell Biol. 8: 34-40.
Warren, G. & Malhotra, V. (1998) The organization of the Golgi apparatus. Curr. Opin. Cell Biol. 10: 493-498.
Wieland, F. T., Gleason, M.L., Serafini, T., Rothman, J.E. (1987) The rate of bulk-flow from the endoplasmic reticulum to the cell surface. Cell, 50:289-300.
Wieland, F.T. & Harter, C. (1999) Mechanisms of vesicle formation: insights from the COP system. Curr. Opin. Cell Biol. 11:440-446.
[1]
Como ejemplo de una proteína de secreción constitutiva ver el artículo de Ureña
& Arribas (2000).