
BIOLOGIA CELULAR |
Las chaperoninas: Máquinas plegadoras de proteínas |
RESUMEN [ABSTRACT]
Las chaperonas moleculares son un amplio grupo de proteínas involucrado en procesos de asistencia en el plegamiento de otras proteínas. Incluida en las chaperonas moleculares hay una familia, la de las chaperoninas o chaperonas moleculares, de peso molecular cercano a 60 kDa (hsp60), que son quizás las mejor caracterizadas. Dos tipos de chaperoninas son las que se conocen, las de origen procariota y pertenecientes a orgánulos citoplasmáticos (grupo I) y las provenientes de arqueobacterias y en el citosol de eucariotas (grupo II). Ambos tipos de chaperoninas comparten una arquitectura muy parecida pero poseen importantes diferencias en el mecanismo de funcionamiento, en los cambios estructurales que se producen durante aquél y en las proteínas a las que asisten en su plegamiento.
El problema del plegamiento de las proteínas
El conocimiento en los primeros años de la década de los cincuenta de que las proteínas están constituidas por una secuencia única (en cada caso) que proviene de la combinación de veinte aminoácidos diferentes hizo que muchos investigadores se hicieran al momento una serie de preguntas: ¿Cómo adquieren su estructura activa las proteínas? ¿Cuáles son los mecanismos que conducen a que una determinada secuencia aminoacídica adopte una estructura con unas propiedades funcionales concretas? La formulación de estos problemas y el intento de responderlos ha dado lugar a una rama de laciencia, hoy en día muy pujante, que se ha denominado "del plegamiento de las proteínas". Mucho se ha investigado desde entonces en este campo y se han producido algunos avances, entre los cuales destacan los descubrimientos realizados por Anfinsen y su euqipo de colaboradores a finales de los años cincuenta, que demostraron que la información necesaria para que una proteína alcance su estructura nativa se encuentra en su propia secuencia. Sin embargo, hoy se sabe que esto no es totalmente cierto, puesto que bajos algunas cisrcunstancias las proteínas no pueden alcanzar su conformación funcional por sí mismas debido a una serie de factores (Figura 1). Uno muy importante y que sólo desde hace poco tiempo ha empezado a tenerse en cuenta, es la alta concentración de macromoléculas existente en la célula (200-400 mg/ml), muy alejada de las condiciones usadas in vitro, y que puede producir interacciones no deseadas entre las proteínas que están siendo sintetizadas. Otro de los problemas con los que se enfrenta en ocasiones la célula son determinadas situaciones de estrés (térmico, salino, ...) que producen la desnaturalización de ciertas proteínas más sensibles a estos estados. Finalmente, las proteínas sintetizadas deben de conservar una conformación desplegada para poder atravesar las membranas de ciertos orgánulos en los que van a actuar.
Las chaperoninas
En muchas de estas ocasiones, las proteínas requieren del auxilio de unas proteínas que empezaron a conocerse a finales de la década de los setenta y que se han denominado chaperonas o acompañantes moleculares (utilizaremos el primer nombre por ser el usado por toda la comunidad internacional). El número de proteínas que se incluye dentro de la familia de las chaperonas es enorme y aumenta de día en día, pero nosotros nos restringiremos a una subfamilia concreta, la de las chaperoninas, que es quizás la más estudiada hasta la fecha.
Las chaperoninas son grandes agregados macromoleculares compuestos por proteínas de un peso molecular cercano a 60 kDa. Todas las chaperoninas conocidas hasta la fecha son oligoméricas y comparten una estructura similar: un cilindro compuesto por uno o dos anillos dispuestos espalda contra espalda (Figura 2). Cada anillo encierra una cavidad, que es el lugar donde se produce el plegamiento de las proteínas. Esta arquitectura es común a todas las chaperoninas, que sólo difieren entre sí en el número de subunidades, iguales o diferentes, de las que se compone cada anillo.

Figura 1. Papel de las chaperoninas en la célula. El papel de las chaperoninas en el plegamiento de proteínas es múltiple. Chaperonas de diversos tipos se unen a los polipéptidos recién sintetizados en los ribosomas (arriba a la izquierda), a proteínas que atraviesan las membranas de diversos orgánulos (arriba en el centro) o a proteínas que se han desnaturalizado debido a cualquier tipo de estrés (arriba a la derecha). Esta unión tiene en un número elevado de casos un papel de protección para evitar que las proteínas alcancen un estado de agregación irreversible. Las chaperonas en general pueden tener un papel activo en el plegamiento de las proteínas, aunque en la mayor parte de los casos parecen transportar los polipéptidos desnaturalizados hasta las chaperoninas, donde son plegados (debajo).
Las chaperoninas de Grupo I
Las chaperoninas más conocidas y estudiadas hasta la fecha son las que se encuentran en eubacterias y orgánulos de organismos eucarióticos (mitocondrias y cloroplastos), que forman parte de las llamadas del Grupo I. Todas las chaperoninas de este grupo tienen una estructura muy parecida, un
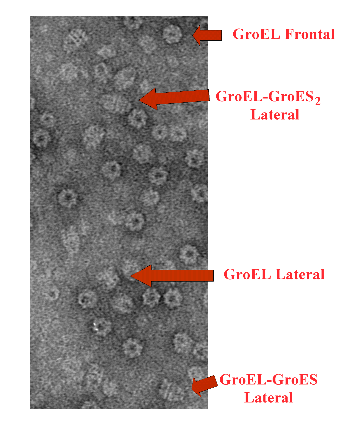
Figura 2. Microscopía electrónica de GroEL. Imagen tomada de un campo de microscopía electrónica de una muestra en la que se ha incubado las chaperoninas GroEL y GroES en presencia de ATP. En el microscopio electrónico GroEL muestra dos tipos de vistas. En una de ellas, la llamada frontal, GroEL tiene la forma de una rosquilla con una cavidad en el centro de la partícula. En la otra vista, denominada lateral, se observa en la partícula cuatro bandas o estrías, de las cuales cada dos corresponden a uno de los dos anillos del oligómero. La vista frontal y lateral de GroEL corresponden al cilindro dispuesto de manera vertical y horizontal, respectivamente, sobre la rejilla del microscopio electrónico. GroES es un pequeño heptámero, apenas visible por si solo en el microscopio electrónico, pero detectable cuando está unido a GroEL. En presencia de nucleótidos de adenina, GroES se une al extremo de uno de los anillos de GroEL en forma de caperuza, tapando de esta manera la cavidad que cada anillo posee. Este complejo GroEL-GroES se denomina asimétrico y tiene la forma de una bala. En condiciones fisiológicas y en presencia de ATP, un oligómero de GroES se une a cada uno de los anillos de GroEL, y da lugar a lo que se ha denominado un complejo GroEL-GroES simétrico, que tiene forma de balón de rugby. Tanto los complejos asimétricos como los simétricos forman parte del ciclo de plegamiento de GroEL.
cilindro constituido por uno odos anillos heptaméricos (Figura 3). La proteína más estudiada dentro de esta familia es GroEL de Escherichia coli. GroEL es una proteína esencial para la viabilidad de la bacteria y experimentos bioquímicos han mostrado que al menos el 10 % de las proteínas celulares interaccionan con esta chaperonina. GroEL se identificó genéticamente
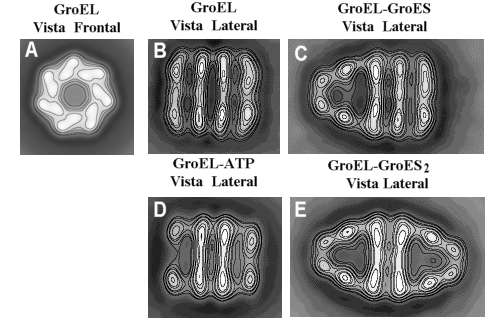
Figura 3. Imágenes medias de GroEL, GroEL-ES y GroEL -ES2. Del promediado de cientos de partículas como las mostradas en la Figura 2 se puede obtener imágenes medias de las distintas confórmeros de GroEL que se pueden obtener durante su ciclo funcional. La adición de ATP induce un cambio conformacional en los dominios apicales de GroEL que se pueden observar en la figura D (apertura de la cavidad), especialmente cuando se la compara con la misma molécula en ausencia de nucleótido (Figura B). La unión de ATP a uno o a los dos anillos permite la unión de GroES a éstos, formando los complejos asimétricos (Figura C) o simétricos (Figura E) que se observan en la Figura 2.
cuando se observó que es indispensable para la morfogénesis de bacteriófagos como l, concretamente para la formación de una subestructura viral, el conector, que une la cabeza con la cola del fago. Sin embargo, durante varios años se estudió la interacción de GroEL con el mecanismo de morfogénesis viral y no su función propia en la bacteria. Cuando se descubrió su homología con otras proteínas de similar peso molecular e involucradas en mecanismos de defensa relacionados con el choque térmico, (Hsp60, proteínas de choque térmico de 60 kDa), GroEL comenzó a ser utilizada por un gran número de laboratorios para caracterizar los mecanismos de asistencia en el plegamiento de proteínas. Los experimentos que se realizaron para probar su función son ahora clásicos. A la manera de los ensayos de Anfinsen, las proteínas a renaturalizar son desnaturalizadas con agentes caotrópicos e incubadas posteriormente en un tampón que incluye GroEL y que además sirve para diluir el agente desnaturalizante. Una vez realizada la incubación con la chaperonina, se ensaya la actividad funcional de la proteína para comprobar el grado de renaturalización alcanzada. Este grupo de ensayo mostró que un gran número de proteínas, muchas de ellas no pertenecientes a E.coli, son capaces de ser replegadas por GroEL. Para ello, y en la mayor parte de los casos, se necesita el concurso de dos cofactores: el primero es un pequeño oligómero formado por siete subunidades de una proteína llamada GroES, que se une a la boca del anillo de GroEL (ambas estructuras son heptaméricas y por lo tanto se ajustan muy bien) para bloquear la cavidad formada por el anillo de GroEL. El segundo de los cofactores es ATP, y su unión y posterior hidrólisis mantendrá en funcionamiento el ciclo funcional de la chaperonina.
Estructuras de GroEL y GroES
De los numerosos estudios realizados por difracción de Rayos X y microscopía electrónica, tenemos una clara de idea de la estructura de GroEL y su cochaperonina GroES, de cómo interaccionan, y de los cambios más drásticos que se producen durante su interacción y durante el ciclo de funcionamiento de GroEL. El monómero de GroEL tiene tres dominios bien definidos (Figura 4): un dominio ecuatorial, que posee la mayor parte de las interacciones que generan el anillo heptamérico y las que mantienen unidos; este dominio también acoge al sitio de unión de ATP, que es el corazón de la maquinaria de GroEL. El segundo dominio es el apical, que se encuentra a la entrada de la cavidad de cada anillo y que posee una agrupación de residuos hidrófobos involucrados en el reconocimiento de los péptidos desnaturalizados que se encuentren en su entorno. Los mismos residuos están involucrados en la unión con la chaperonina GroES, y este hecho es de capital importancia en el mecanismo de funcionamiento de GroEL. Finalmente, existe un tercer dominio, el intermedio, que ejerce de transmisor de las señales que se producen entre los dominios ecuatorial y apical y funciona como una bisagra para trasladar los grandes cambios conformacionales que, producto de la unión de ATP y GroES, ocurren en el dominio apical.
La estructura de GroES es mucho menos compleja (Figura 4). Cada monómero de esta cochaperonina tiene una estructura de barril de láminas b. La mayor parte del monómero está estructurado excepto un gran lazo que se encuentra desordenado, pero que cuando interacciona con el dominio apical de GroEL, se une fuertemente a éste y genera en él un gran cambio conformacional. El heptámero de GroES, dependiendo del momento del ciclo funcional de GroEL en que se encuentre, puede unirse a uno de los anillos de GroEL o a los dos, formando lo que se ha denominado respectivamente, complejos asimétricos o simétricos de GroEL y GroES (Figura 3).
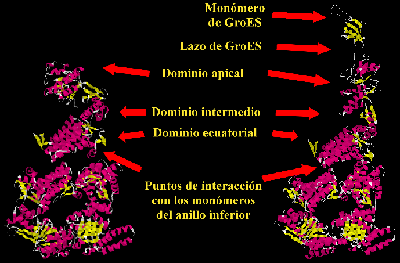
Figura 4. Estructura atómica de los monómeros de GroEL y de GroES. En ambas imágenes se observa la estructura atómica de un monómero de GroEL correspondiente al anillo superior de GroEL, enfrentado a los dos monómeros del anillo inferior con los que interacciona. En la imagen de la izquierda se representa la conformación abierta y susceptible de aceptar polipéptidos desnaturalizados, en la que el dominio apical del monómero superior se encuentra en la posición basal. La unión de ATP en el sitio de unión de este nucleótido (situado en el dominio ecuatorial) y la posterior unión de la cochaperonina GroES (de la que sólo se muestra un monómero en la imagen de la derecha) transmite al dominio apical a través del dominio intermedio una señal para que en aquél se genere un gran cambio conformacional de tal manera que ahora se eleva y apunta hacia arriba, lo que produce un considerable aumento de la cavidad de GroEL.
Mecanismo funcional de GroEL
Durante los últimos años se han realizado multitud de experimentos de carácter bioquímico y biofísico que han permitido tener una idea bastante clara de cual es el mecanismo de funcionamiento de GroEL y por ende, de todas las chaperoninas del grupo I (Figura 5):
1)Los residuos hidrofóbicos que forman un anillo alrededor de la boca de la cavidad de cada anillo funcionan como un imán que reconoce y une todos los residuos hidrofóbicos que se encuentran a su alrededor. Si se tiene en cuenta que las proteínas celulares que se encuentran en solución mantienen los residuos hidrofóbicos en el interior de su estructura, la exposición de éstos significa que esas proteínas no se encuentran plegadas correctamente. El mecanismo de reconocimiento utilizado por GroEL es por lo tanto muy simple y muy inespecífico. De ahí el amplio rango de proteínas a las que asiste en su plegamiento.
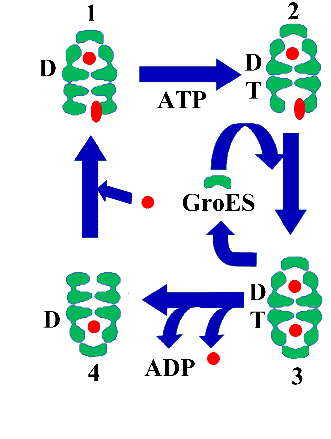
Figura 5. Mecanismo de funcionamiento de GroEL. En el esquema se observa el ciclo alternado que poseen los dos anillos de GroEL. Así, mientras que el anillo superior ya posee en su cavidad una molécula de polipéptido dispuesta a plegarse por si mismo, el anillo inferior ha reconocido y unido otro polipéptido desplegado (posición 1). La unión de ATP a este anillo (posición 2) permite el cambio conformacional de los dominios apicales que dará lugar a la unión de GroES (posición 3) y a la señal de liberación de GroES del anillo superior, con la consiguiente liberación del polipéptido ocluido en este anillo (posición 4). El anillo superior, en este punto, se encuentra en la misma conformación que el anillo inferior en la posición 1.
2) ATP se une a su sitio de unión, en el dominio ecuatorial del monómero de GroEL. La unión de ATP desencadena una cadena de señales en varias direcciones:
Existe una cooperatividad positiva entre los monómeros de un mismo anillo, de manera que el resto de los monómeros de ese anillo unen ATP inmediatamente.
Se produce una cooperatividad negativa entre las sub unidades de los dos anillos, de manera que una vez que una molécula de ATP se une a un monómero de los anillos y desencadena la unión inmediata de moléculas de ATP a las otras seis sub unidades del mismo anillo, se impide la unión de ATP a las siete sub unidades del otro anillo.
Se envía una señal desde el dominio ecuatorial de cada sub unidad que ha unido ATP, a través del dominio intermedio hasta el dominio apical, en el que se produce un cambio conformacional (elevación de la punta del dominio apical) que permite la unión de la cochaperonina GroES.
3) La unión del oligómero de GroES induce un levantamiento aún mayor de las puntas de los dominios apicales de GroEL y por lo tanto un aumento de la cavidad dentro del anillo. Esta unión genera la formación del complejo asimétrico descrito en la Figura 3. Por otra parte, la unión de GroES sella la cavidad y es en ésta donde se va producir el fenómeno más importante de todo el ciclo. Los residuos hidrofóbicos que hasta ahora interaccionaban con el polipéptido desnaturalizado lo hacen ahora con el lazo desordenado de GroES. El cambio conformacional promovido por la unión a la cochaperonina hace que todos los residuos hidrofóbicos y cargados que se exponían en la superficie interior de la cavidad, ahora desaparecen y dejan paso a residuos hidrofílicos. El resultado de todo ello es que el polipéptido, que había sido atrapado anteriormente por la boca del anillo, es encerrado en éste y su salida de la cavidad es imposibilitada por la cochaperonina que tapa la boca de ésta. En este estado, y libre de las interacciones no deseadas que le acechan en el exterior, (interacciones con los residuos hidrofóbicos de otras proteínas que dan lugar a agregados irreversibles), y en una cavidad que se ha hecho mucho más grande por efecto de los cambios conformacionales que han sufrido los dominios apicales, puede el polipéptido intentar plegarse por sí mismo utilizando la información codificada en su secuencia primaria. Algunos experimentos sugieren que durante el cambio conformacional inducido por la unión de ATP y GroES, los dominios apicales tiran del sustrato de manera que lo despliegan antes de liberarlo en la cavidad.
4)Las moléculas de ATP unidas en todas las sub unidades del anillo se hidrolizan. La hidrólisis del nucleótido, además de relajar la unión de GroES a GroEL en ese anillo, envía una señal al otro anillo para indicarle que ya puede unir ATP. Esta unión se produce, con los consiguientes cambios indicados anteriormente en el punto 2). La unión de ATP permite también que otra molécula de GroES se una a este segundo anillo, formando un complejo simétrico, con una cochaperonina unida a cada anillo de GroEL, tal y como se describe en la Figura 3.
5)La formación del complejo simétrico no es muy estable, porque la unión de GroES al segundo anillo induce la liberación de la cochaperonina del primer anillo y la salida al exterior del polipéptido encerrado en aquel. El primer anillo está dispuesto para recibir otro polipéptido desnaturalizado que se encuentre en su entorno. Los pasos 4) y 5) se reproducen ahora en el segundo anillo.
Podemos imaginarnos pues al oligómero de GroEL como un motor de dos cilindros (los dos anillos), en el que cada uno de ellos se encuentra en cada momento en posiciones diferentes del ciclo. Mientras un anillo está liberando la cochaperonina y por ende el polipéptido encerrado, el otro está uniendo otra cochaperonina. Los dos anillos funcionan de manera alternada, ya que cada uno de ellos controla al otro mediante la transmisión de señalesya mencionada, señales que van desde los dominios ecuatoriales de las sub unidades de un anillo al del otro, y viceversa. La unión y posterior hidrólisis del ATP funciona como el combustible de los cilindros, lo que permite que el ciclo se mantenga funcionando indefinidamente.
Significado fisiológico del papel de las chaperoninas
El gasto energético que se produce durante el ciclo de GroEL no incide directamente sobre la función de la chaperonina en la asistencia en el plegamiento de proteínas, sino que tan solo sirve para mantener activo el ciclo. En este sentido debe de quedar claro que GroEL y las chaperoninas en general no son enzimas, pues no modifican covalentemente al sustrato con el que interaccionan, sino que permiten que dentro de su cavidad el polipéptido desnaturalizado pueda alcanzar libremente su estructura nativa. Esta puede ser alcanzada por parte del polipéptido dentro de la cavidad, o fuera de ella después de su liberación. Puede también que el polipéptido desnaturalizado se mantenga en esta conformación después de una primera interacción con la chaperonina, en cuyo caso puede ser reconocido otra vez por ésta, con lo que vuelve a intentar su plegamiento. Hay varios estudios bioquímicos que demuestran que distintas proteínas tienen estadísticamente una media de interacciones con GroEL diferente. El mecanismo de GroEL es ineficiente (genera un gasto continuo de ATP independientemente o no de que haya moléculas de sustrato desnaturalizado en la cavidad de la chaperonina; no siempre repliega a la proteína en un solo ciclo de interacción) pero en contrapartida funciona para un rango muy grande de sustratos (en la práctica, y excepto en el caso de ciertas proteínas, puede ayudar a replegar a cualquier proteína con un peso molecular menor de 50 kDa).
Siendo GroEL una máquina molecular muy compleja, posee algunos mecanismos aun más sofisticados, como los cambios que se producen en su funcionamiento durante periodos de choque térmico. GroEL y la mayor parte de las chaperoninas del grupo I son proteínas de choque térmico y ven inducidas su producción (hasta diez veces más) durante los momentos en que aquél se produce para tratar de lidiar con el mayor número de proteínas que se desnaturalizan por el aumento de temperatura. Sin embargo, no parece tener sentido que estas proteínas sean renaturalizadas para que vuelvan a ser desnaturalizadas si el choque térmico es prolongado. Pues bien, ahora se conoce que al aumentar la temperatura, la unión de la cochaperonina GroES a GroEL es más fuerte, de tal manera que aquélla tarda más en liberarse y mantiene por lo tanto al polipéptido encerrado en la cavidad más tiempo. Este mecanismo diferente se debe a un cambio conformacional inducido por la temperatura, que altera la señal de liberación de la cochaperonina que envía un anillo al otro. Este fenómeno y quizás algún otro que resta por caracterizar explicaría por qué las chaperoninas están constituidas por dos anillos, cuando cada uno de ellos por separado podría formar una unidad funcional en la asistencia al plegamiento.
Chaperoninas del Grupo II
El otro tipo de chaperoninas, aquéllas que se encuentran en arqueobacterias y en el citoplasma de organismos eucarióticos, es mucho menos conocido, y sólo en los últimos años se han comenzado a realizar caracterizaciones bioquímicas y estructurales de éstas. Por de pronto, y a diferencia de las chaperoninas del grupo I, las de grupo II son muy variables en su grado de oligomerización y en el número de sub unidades distintas que componen el oligómero. Así, la estructura de doble anillo puede constituida por dos anillos octaméricos ó nonaméricos, y los anillos formados por uno, dos, tres o incluso ocho proteínas diferentes. Este caso último es el de la chaperonina citoplasmática de eucariotas ó CCT, que es quizás la chaperonina más interesante por una serie de razones: además de poseer ocho sub unidades diferentes (aunque homólogas entre sí) en cada uno de los dos anillos octaméricos, asiste específicamente al plegamiento de actinas y tubulinas, dos proteínas citoesqueléticas de vital importancia para la célula.
Los estudios estructurales realizados con CCT y alguna chaperonina de arqueobacterias muestran que éstas poseen los mismos dominios (ecuatorial, intermedio y apical) que GroEL, así como sus dos mismas conformaciones básicas (Figura 6). Se ha caracterizado una conformación abierta, que se producen en ausencia de ATP, y en la que la cavidad se ofrece accesible para la interacción con el sustrato. La unión de ATP genera la conformación cerrada, pero a diferencia de GroEL, el cierre de la cavidad no se produce por la unión de una cochaperonina, ya que las chaperoninas del grupo II no poseen cochaperoninas, sino que un dominio extra que se encuentra en los dominios apicales cierra la cavidad al producirse el cambio conformacional. El sustrato que se ha unido previamente a la chaperonina queda así cerrado en su cavidad.
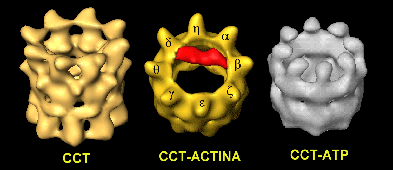
Figura 6. Estructuras de CCT, CCT-ATP y CCT-actina. . Las estructuras tridimensionales de los distintos confórmeros de la chaperonina CCT que se muestran aquí se han obtenido mediante microscopía electrónica y procesamiento de imagen, y revelan la típica estructura de dos anillos común a todas las chaperoninas, con la particularidad que en este caso el oligómero está constituido por dos octámeros generados por ocho distintas sub unidades (imagen izquierda). La adición de ATP (imagen derecha) genera un cambio en los dominios apicales de tal manera que éstos apuntan hacia el interior y cierran la cavidad (aunque la baja resolución de la reconstrucción no permite visualizar el cierre completo de aquélla) imitando al confórmero de GroEL en el que se encuentra unida la cochaperonina GroES. La adición a la estructura libre de actina desnaturalizada permite la unión de ésta de dos maneras específicas: en uno de ellos un brazo de actina (coloreada en rojo) se une al dominio apical de la subunidad y otro al de la (imagen central), y en otro caso un brazo de actina se une a la subunidad y otro a la no representado aquí.
¿Qué tipo de interacción se produce entre la chaperonina y el sustrato? Muy poco se conoce respecto a los sustratos de las chaperoninas de arqueobacterias, y la información que se ha obtenido hasta la fecha proviene de los estudios realizados con CCT. La interacción entre CCT y el sustrato parece ser de distinta naturaleza a la que se establece entre GroEL y las proteínas a las que asiste en su plegamiento. Mientras GroEL reconoce residuos hidrofóbicos que se encuentran expuestos al azar en el solvente, actina se une a CCT a través de interacciones entre residuos específicos de CCT y actina. Estas interacciones se producen con sub unidades concretas de CCT (Figura 6), mientras que en el caso de GroEL, los polipéptidos que se unen a esta chaperonina lo hacen al azar a cualquiera de las siete sub unidades. Otra de las diferencias importantes es que los sustratos con los que GroEL interacciona están parcial o totalmente desnaturalizados, mientras que parece que actina se une a CCT en una conformación cuasi nativa a la que sólo le falta un pequeño pero importante paso para alcanzar la conformación nativa. Ese paso sólo se lo puede proveer CCT, de tal manera que ciertas mutaciones en esta chaperonina o su ausencia provocan la muerte celular. La especificidad de CCT sobre sus sustratos (actina y tubulina principalmente) y la de éstos por CCT se refuerza por el hecho de que GroEL interacciona promiscuamente con cualquier proteína, incluso actina y tubulina en un estado desnaturalizado, pero no es capaz de plegarlas.
Todo parece indicar que CCT apareció en la naturaleza a la vez que los organismos eucariotas (ya que está presente sólo en ellos), como evolución de una chaperonina primigenia y en respuesta a los problemas de plegamiento que plantea la tubulina, proteína que apareció en los organismos eucariotas como una evolución de la más primitiva FtsZ, presente en procariotas. La tubulina es una proteína muy compleja, que forma estructuras enormes llamadas microtúbulos como consecuencia de complejos fenómenos de polimerización. La formación de los microtúbulos tiene que ver con peculiaridades en la estructura de tubulina, y es posible que la aparición de aquéllas haya dado lugar a la evolución de una chaperonina específica para lidiar con un problema muy puntual pero vital a la célula como es el plegamiento de tubulina (y el de actina). El hecho de que FtsZ no interaccione con CCT y su plegamiento se produzca por si sola o con la ayuda de GroEL parece reforzar la hipótesis aquí apuntada.
En resumen, los resultados obtenidos hasta la fecha nos muestran a las
chaperoninas como unas máquinas moleculares con una arquitectura
ideada por la naturaleza para tratar un problema general, el del plegamiento
de proteínas, utilizando para ello un mecanismo que aunque ineficiente
desde el punto de vista energético se adapta a un gran número
de proteínas. Durante la evolución natural de los organismos,
esa misma arquitectura ha sido utilizada por otra chaperonina para tratar
de forma específica y muy eficiente el problema del plegamiento
de dos proteínas citoesqueléticas vitales para la célula
como son la actina y la tubulina.
Referencias
Anfinsen, C.B. (1973). Principles that govern the folding of protein chains. Science 181, 223-230.
Braig, K. Otwinowski, Z., Hegde, R., Boisvert, D.C., Joachimiak, A., Horwich, A.L. & Sigler, P.B. (1994) The crystal structure of the bacterial chaperonin GroEL at 2.8 Å. Nature 371, 578-586.
Bukau, B. & Horwich, A.L. (1998) The hsp70 and hsp60 chaperone machines. Cell 92, 351-366.
Ditzel, L., Löwe, J., Stock, D., Stetter, K.O., Huber, H., Huber, R. & Steinbacher, S. (1998) Crystal structure of the thermosome, the archaeal chaperonin and homologue of CCT. Cell 93, 125-138.
Ellis, R.J. (1994). Opening and closing the Anfinsen cage. Curr. Biol. 4, 633-635.
Ellis, R.J., and Hartl, F.U. (1999). Principles of protein folding in the cellular environment. Curr. Opinin. Struct. Biol. 9, 102-110.
Houry W. A., Frishman D., Eckerskorn C., Lottspeich F., and Hartl F. U. (1999) Identification of in vivo substrates of the chaperonin GroEL. Nature, 402, 147-154.
Llorca, O., Galán, A., Carrascosa, J.L., Muga, A. & Valpuesta, J. M. (1998) GroEL under heat shock: switching from a folding to a storing function. Journal of Biological Chemistry 273, 32587-32594.
Llorca, O., McCormack, E. Hynes, G., Grantham, J., Cordell, J., Carrascosa, J.L., Willison, K.R., Fernández, J.J. & Valpuesta, (1999) J.M. Eukaryotic type II chaperonin CCT interacts with actin through specific subunits. Nature 402, 693-696.
Llorca, O., Smyth, M.G., Carrascosa, J.L., Willison, K.R., Radermacher, M., Steinbacher, S. & Valpuesta, J.M. (1999) 3D reconstruction of the ATP-bound form of CCT reveals the asymmetric folding conformation of a type II chaperonin. Nat. Struct. Biol. 6, 639-642.
Willison, K.R. Composition and function of the eukaryotic cytosolic chaperonin-containing TCP1 in Molecular Chaperones and Folding Catalysts (ed. Bernd Bukau) 555-571 (Harwood Academic Publishers, Amsterdam, 1999).