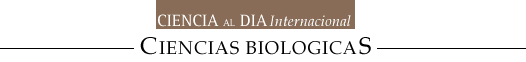
NEUROCIENCIAS |
La acción neuromoduladora de la dopamina sobre los ganglios basales |
|
© José Bargas, Elvira Galarraga, 2000
jbargas@ifisiol.unam.mx, Instituto de Fisiología Celular de la UNAM, México |
RESUMEN [ABSTRACT]
Los acúmulos de neuronas cerebrales no corticales conocidos como "ganglios basales" (GB) intervienen en la regulación de las funciones motoras. Las funciones motoras son comandadas por la actividad neural de las cortezas (Cx) motora y premotora. Los comandos corticales se dirigen a la médula espinal donde están las motoneuronas encargadas de activar a los músculos involucrados en el movimiento. Los GB regulan estos comandos o "programas" de activación motora generados por la Cx mientras están en marcha. Los GB reciben entradas de la Cx sensorial que ayudan a corregir y afinar los programas motores. Como la mayoría de los circuitos cerebrales, los GB utilizan la neurotransmisión rápida, asociada a receptores-canal activados por ligando, y los neurotransmisores glutamato (excitador) y ácido gama-aminobutírico (GABA) (inhibidor), para conformar sus circuitos. Sin embargo, la neurotransmisión rápida está regulada por la llamada neurotransmisión lenta o moduladora, asociada a receptores que activan cadenas de señalización intracelular (e.g., proteínas G). Acaso, el neurotransmisor modulador más importante de los GB es la dopamina (DA). Sin DA, el circuito de los GB sufre un colapso total y deja de funcionar. Esto constituye un ejemplo dramático de la importancia de la neuromodulación. Esto es lo que parece suceder en la enfermedad de Parkinson, que es quizás, la primera patología reconocida como desorden de la neuromodulación. El tratamiento de esta enfermedad se basa en la terapia restitutiva con análogos o precursores del neuromodulador DA. Asimismo, diversas enfermedades motoras se conciben ahora como desbalances neuromodulatorios en el circuito de los GB (coreas, distonias, etc.). En el presente trabajo revisaremos el circuito de los GB y las acciones moduladoras de la DA.
Breve introducción al funcionamiento del sistema motor
Los grupos de neuronas centrales que controlan a las motoneuronas que generan el movimiento están en el tallo cerebral (o TC - bulbo, puente, mesencéfalo y cerebelo) y en el cerebro (corteza cerebral -Cx- y ganglios basales -GB). Para que un movimiento sea ejecutado por una extremidad se requiere: a) activar los músculos agonistas que ejecutan el movimiento, e.g., flexores, b) inhibir los músculos antagonistas que se oponen a dicho movimiento, e.g., extensores, y c) activar ciertos músculos posturales o antigravitatorios para preservar el centro de gravedad del cuerpo, y así mantener el equilibrio mientras el movimiento se efectúa. Así, el correlato neural de todo movimiento requiere que las neuronas de unos músculos se activen y las de otros se inhiban. Los grupos de neuronas que mantienen el tono muscular y la postura están situados en el TC, en la región que se denomina "formación reticular", que recibe entradas aferentes sensoriales de la médula espinal y del aparato vestibular. Estas neuronas están todo el tiempo activas, pero reciben influencias inhibidoras de los GB, del cerebelo y de la Cx, que disminuyen su activación de acuerdo a la demanda. Si estas vías inhibidoras se lesionan, la substancia reticular se "libera" generando una sobreactivación motora que causa rigidez, parálisis espástica y aumento generalizado del tono muscular (Suszkiw, 1996). Esto sugiere que el llamado "reposo" (relajación y ausencia de movimientos visibles) es un proceso activo generado por el balance entre la actividad tónica de las neuronas del tallo cerebral y la actividad inhibidora producida por la masiva innervación cerebral (Cx, GB) sobre las interneuronas inhibidoras del tallo.
Cuando se ejecuta un movimiento no se parte de cero para comenzar a activar a las neuronas motoras involucradas (tal y como lo hacen algunos prototipos de robots), sino que siempre hay una actividad basal sobre la cual se sobreponen, de manera fásica y secuencial, aumentos o disminuciones de la actividad. El único momento en que la actividad basal disminuye casi por completo es durante el sueño paradójico. La Figura 1 es un esquema general del circuito del sistema motor, incluyendo el lugar que corresponde a los GB.
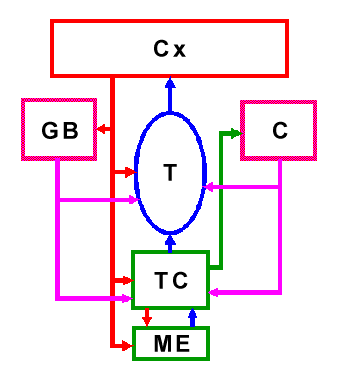
Figura 1. Lugar de los ganglios basales en el sistema motor. Cx: corteza cerebral, GB: ganglios basales, T: tálamo, C: cerebelo, TC: tallo cerebral, ME: médula espinal. Las vías ascendentes sensoriales (en azul) llegan a la Cx haciendo sinapsis en T. El cerebelo recibe aferentes sensoriales del TC, provenientes de la ME y el núcleo vestibular (en verde). La Cx también se comunica con el C a través del TC (en verde). La Cx genera las vías descendentes que llevan los comandos motores (en rojo). Algunas llegan directamente a la ME, muchas llegan al TC para regular múltiples vías descendentes (rubroespinal, vestíbulo espinal, tecto espinal, retículo espinal) junto con la regulación de los GB y el C (en magenta). La Cx se comunica directamente con los GB llevando información sensorial y motora (en rojo). Los GB modifican la información de Cx y la envían al T y TC (en magenta). El T regresa la información modificada a la Cx motora y premotora.
Los ganglios basales
Los GB comprenden grupos neuronales de entrada, como el neoestriado (o NE, que comprende a los núcleos caudado y putamen); núcleos de salida como el globo pálido interno (GPi) y la sustancia negra reticulada (SNr) y núcleos intrínsecos como la sustancia negra compacta (SNc), el globo pálido externo (GPe) y el núcleo subtalámico (NST). En la figura 2 se esquematizan los núcleos que forman parte de los GB.
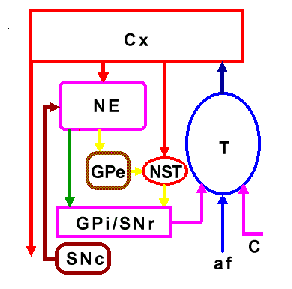
Figura 2. Componentes de los Ganglios Basales (GB). Cx: corteza cerebral, NE: neoestriado (núcleos caudado y putamen), GPi: globo pálido interno, SNr: substancia negra reticulada, GPe: globo pálido externo, SNc: substancia negra compacta, origen de la aferencia dopaminérgica al NE (en café), NST: núcleo subtalámico. C: cerebelo, af: aferentes. Red directa en verde. Red indirecta en amarillo. GPi/SNr: núcleos de salida. Proyecciones corticales en rojo. Proyecciones de los núcleos de salida al T en magenta. Se usa la palabra "red", en vez de "vía", pues como se explica en el texto, se trata de conectar grupos neuronales que no necesariamente se activan de manera igual o al unísono. La activación diferencial de estos grupos neuronales (como la función de contraste en un monitor de TV) es lo que es facilitado por la dopamina.
El NE recibe aferentes de toda la corteza (sensorial, motora y de asociación). Esto es, recibe copias de perceptos y de programas motores en ejecución. Grupos de neuronas piramidales arregladas en columnas disparan sincrónicamente y en secuencia para activar a los músculos que ejecutan un determinado movimiento. Hay grupos de neuronas dedicados a cada músculo del cuerpo. Estas neuronas piramidales excitan de manera simultánea a las neuronas de los GB (Fig. 2). La conexión entre Cx y NE utiliza neurotransmisión rápida, o sea, las aferentes corticales al NE son glutamatérgicas y activan receptores-canal de los tipos AMPA y NMDA en las neuronas post-sinápticas (ver Glosario). El 95% de las neuronas neoestriatales post-sinápticas son de proyección y GABAérgicas (llamadas "espinosas medianas" debido a la cantidad de espinas dendríticas que poseen), y responden a la activación cortical con trenes de potenciales de acción que anteceden en decenas de milisegundos a la activación muscular (Wilson, 1998). La micro-estimulación de pequeños grupos de neuronas neoestriatales activa grupos musculares alrededor de una articulación (Alexander, 1987). Por lo anterior, la actividad de las neuronas espinosas es considerada "la salida funcional" del NE y constituye el punto de arranque de todos los modelos sistémicos y fisiopatológicos de los GB (e.g. Alexander, 1995; Mink, 1996). .
Redes directa e indirecta.
Las neuronas del NE proyectan hacia afuera del núcleo de dos maneras: Un grupo de neuronas proyecta directamente sobre los núcleos de salida, el GPi y la SNr (flecha verde en la Fig. 2), mientras que otro grupo proyecta sobre el GPe (flecha amarilla en la Fig. 2). El primer grupo inicia la llamada "red directa", y el segundo origina la llamada "red indirecta".
Los núcleos de salida, GPi y SNr (Fig. 2), innervan a los núcleos basales del tálamo (T) (flecha magenta en la Fig. 2). De los núcleos talámicos surgen circuitos tálamo-corticales que modifican la actividad de salida de las cortezas motora y premotora (flecha azul de T a Cx en la Fig. 2). Así, el circuito se inicia y termina en la Cx. Sin embargo, la actividad neural es regresada a la Cx después de ser contrastada con entradas sensoriales y ser transformada por las redes "directa" e "indirecta" de los GB. La investigación actual trata de entender qué "le hacen" las redes directa e indirecta de los GB a los comandos motores de la corteza. Una de las hipótesis postula que los GB ayudan a separar funcionalmente a las poblaciones de neuronas que activan músculos agonistas de las que activan músculos antagonistas (Mink, 1996). Esto tiene que hacerse en rápida sucesión pues un programa motor está compuesto de una secuencia de comandos que activan y desactivan a los grupos musculares involucrados siguiendo un patrón temporal. Los músculos antagonistas para un movimiento pueden ser sinergistas para otro movimiento. Así, muchos programas motores sólo son aprendidos después de muchos ensayos y práctica, por lo que los GB están involucrados en el condicionamiento, la memoria y el aprendizaje (Schultz, 1998).
Las neuronas de los núcleos de salida, el GPi y la SNr, son GABAérgicas inhibitorias. Su actividad es tónica, i.e., disparan potenciales de acción todo el tiempo a frecuencias de entre 20 y 50 Hz, produciendo una inhibición sostenida de las neuronas de los circuitos motores tálamo-corticales. GPi y SNr poseen una representación topográfica de todos los músculos del cuerpo. La actividad tónica de estos núcleos mantiene inhibida la activación de grandes grupos musculares y se opone a la activación generalizada provocada por la actividad de la formación reticular del TC. Para que pueda realizarse la activación de algunos músculos, e.g., para realizar un movimiento voluntario, hay que inhibir selectivamente a las neuronas inhibitorias del GPi o SNr que impiden los movimientos de esos músculos (flecha verde en Fig. 2) (Alexander, 1995).
Las neuronas de GPi y SNr pueden inhibirse selectivamente cuando reciben andanadas de potenciales de acción enviados por las neuronas GABAérgicas neoestriatales de la red directa. Estas ráfagas de potenciales de acción, como ya vimos, están comandadas por la Cx (en rojo, Fig. 2) (Wilson, 1998). La conexión involucra neurotransmisión rápida inhibitoria utilizando el receptor canal GABAA. Así, dos grupos de neuronas inhibitorias en serie, uno en el NE y otro en el GPi/SNr (Fig. 2), permiten realizar una activación neta y focalizada de los núcleos talámo-corticales mediante "desinhibición". Esta desinhibición facilita los movimientos voluntarios originados en la Cx (Alexander, 1995).
Papel de la dopamina sobre las neuronas de la red directa
El neurotransmisor DA, liberado por las neuronas de la SNc que innerva al NE, activa receptores para la dopamina (dopaminérgicos), principalmente de un tipo particular llamado D1, en las neuronas neoestriatales de la red directa. Estos receptores están acoplados a una cadena de señalización intracelular. Su activación hace que aumente la excitación que ejerce la actividad cortical sobre las neuronas espinosas del NE (Gonon, 1997); ésto aumenta la activación (desinhibición) de las neuronas tálamo-corticales que activan a los músculos agonistas, y por lo tanto, facilita los movimientos (Alexander, 1995).
El efecto "procurador" de DA sobre la excitabilidad del NE se debe a la facilitación de corrientes persistentes de calcio en las neuronas espinosas (Surmeier et al., 1995; Hernández-López et al., 1997) y a la facilitación de corrientes sinápticas de tipo NMDA (Cepeda y Levine, 1998). Ambos efectos llevan a un aumento en la actividad de las neuronas espinosas (ver Fig. 3). Sin embargo, esta acción facilitadora de la dopamina sólo se presenta si las neuronas están siendo despolarizadas por los comandos corticales. Sólo así se alcanzan los voltajes de activación para los canales iónicos involucrados (de calcio tipo L y NMDA). Sin embargo, la DA tiene otros efectos. Que se presente la acción facilitadora o inhibidora depende del potencial de membrana de la neurona post-sináptica. La DA es excitadora a potenciales de membrana despolarizados, presentes cuando la célula está siendo activada, y es inhibidora, aumentando el umbral de disparo, a potenciales de membrana hiperpolarizados, presentes cuando la célula está en reposo o inhibida (Fig. 3). La DA es inhibidora a potenciales de membrana cercanos a los de reposo debido a que también aumenta la activación de canales de potasio (rectificadores de entrada) a potenciales de membrana negativos (Pacheco-Cano et al., 1995) (Fig. 3). Esto aumenta el umbral de disparo neuronal, lo que hace que sea más difícil activar una neurona. Ambas acciones de la DA, facilitadora e inhibidora, pueden observarse en la misma neurona, y ambas se deben a la activación del receptor D1 (Fig. 3). Lo que nos lleva a ejemplificar una diferencia entre la neurotransmisión rápida y la neuromodulación: mientras que la neurotransmsión rápida sólo activa uno o dos tipos de receptor-canal sobre la neurona post-sináptica, la neuromodulación puede modificar la actividad de varios tipos de canal simultáneamente, provocando así, modificaciones complejas en la excitabilidad; que pueden depender del potencial de membrana. Esto se debe a que el receptor puede acoplar sus cadenas de señalización con varios efectores (e.g., canales iónicos) al mismo tiempo y que los canales iónicos dependientes de voltaje poseen diferentes voltajes de activación.
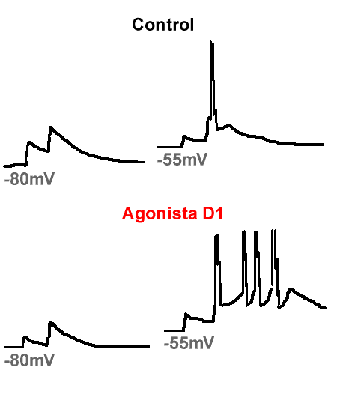
Figura 3. Acciones excitadora e inhibidora de la dopamina (DA). Un par de potenciales sinápticos excitatorios es provocado en una neurona neoestriatal de proyección estimulando las aferentes corticales. En la columna de la izquierda los potenciales sinápticos son provocados desde un potencial de membrana de -80 mV. En la columna de la derecha la misma estimulación es usada para provocar los potenciales sinápticos desde un potencial de membrana de -55 mV. Los potenciales sinápticos son de mayor amplitud a -80 mV pues este potencial está más alejado del potencial de inversión. Sin embargo, no llegan al umbral de disparo. En presencia de una agonista del receptor dopaminérgico tipo D1 (SKF 81297) los potenciales se reducen en amplitud y se alejan aun más del umbral de disparo pues la activación del receptor D1 abre canales de potasio (rectificador de corriente entrante del tipo IRK) y disminuye la resistencia de entrada. Cuando se provocan a -55 mV, uno de los potenciales sinápticos llega al umbral de disparo (el potencial de acción está truncado). En presencia del agonista del receptor D1, el potencial sináptico genera una despolarización prolongada que puede sostener una ráfaga de potenciales de acción. La despolarización prolongada está ocasionada por la facilitación simultánea de las corrientes tipo NMDA y L (no publicado previamente pero ver: Cepeda y Levine, 1998; Galarraga et al., 1997; Hernández-López et al., 1997).
El que la DA sólo facilite a las neuronas activadas por la Cx, mientras que ayude a inhibir a las neuronas no activadas, hace que se separen mejor las neuronas que activan a los músculos agonistas de las neuronas que activan a los músculos antagonistas. Esta separación es necesaria para que las secuencias de activación y desactivación muscular se sucedan tersamente y no de manera brusca o escalonada. Así, cuando esta separación es deficiente, hay co-activación de músculos agonistas y antagonistas; lo que provoca rigidez y aun oposición de las extremidades a ser desplazadas pasivamente (signo de "rueda dentada"). Todos estos signos son característicos de la enfermedad de Parkinson. En la enfermedad de Parkinson hay degeneración de las neuronas dopaminérgicas y depleción de la dopamina neoestriatal.
La acción de la DA se suma a otras acciones del circuito de los GB encaminadas a separar la activación de músculos agonistas y antagonistas. Otro mecanismo encargado de esta separación es la inhibición colateral recíproca: Las colaterales axónicas de las neuronas neoestriatales activadas, inhiben a las neuronas espinosas vecinas, ya que las neuronas espinosas se conectan entre sí mediante sinapsis GABAérgicas (este mecanismo está a discusión, véase: Wickens y Oorschot, 2000). Se sabe que los receptores dopaminérgicos del tipo D1, también están situados en las terminales axónicas de las neuronas de la red directa, por lo que su activación aumentaría la liberación de GABA de estas terminales, reforzando así la inhibición lateral (Aceves et al., 1995). Así, la DA amplificaría la inhibición lateral actuando presinápticamente, ayudando a separar mejor las poblaciones neuronales activadas de las no activadas. Actualmente realizamos experimentos para descartar o reforzar esta hipótesis.
Por otro lado, la DA también cuenta con receptores presinápticos en las terminales de algunas aferentes glutamatérgicas que llegan al NE. Se piensa que las acciones pre-sinápticas de la DA a este nivel, ayudan a "seleccionar" ciertas aferentes o entradas excitatorias, mientras otras son bloqueadas (Flores-Hernández et al., 1997).
Un potencial de membrana muy polarizado (< -80 mV), el carácter silente (no generan potenciales de acción ni potenciales marcapaso espontáneamente), la fuerte rectificación por corrientes de potasio facilitadas por la dopamina, y la inhibición lateral, son todas características fisiológicas que hacen muy difícil propagar la actividad de las neuronas espinosas del NE. A esto debemos de agregar que cada aferente cortical produce muy pocos botones sinápticos sobre una neurona espinosa ;: de 1 a 2 en promedio. Así, el NE es clasificado como un núcleo "no epileptogénico", o que no genera crisis epilépticas. Un comando cortical debe sobrepasar cierta intensidad para pasar el "filtro" del NE. En términos de fisiología cortical, se tienen que sincronizar varias columnas de neuronas piramidales para poder activar a las neuronas espinosas. Ahora bien, cuando el comando cortical es suficientemente intenso y sincrónico como para generar un tren de potenciales de acción en una neurona espinosa, entonces la DA facilita y prolonga la respuesta en virtud de las acciones moduladoras que hemos descrito. En otras palabras, una vez que el estímulo pasa el filtro, la dopamina no deja que decaiga la señal si no que la amplifica. Esto "afina" el comando motor mejorando la proporción señal/ruido.
Cuando el "filtro" neoestriatal se daña, i.e., cuando las neuronas neoestriatales se lesionan (por ejemplo en el caso de la enfermedad de Huntington), aparecen movimientos involuntarios que interrumpen y estorban la ejecución de los movimientos voluntarios ("ruido") produciendo incapacidad (e.g., síndromes "hipercinéticos" tales como tics, coreas, distonias y atetosis).
La red indirecta
La red indirecta también posee dos neuronas inhibitorias en serie (flechas amarillas en la Fig. 2). Las neuronas neoestriatales de esta red inhiben a las neuronas del GPe cuando reciben un comando cortical. Las neuronas del GPe son GABAérgicas, disparan de manera tónica, e inhiben a su vez, a las neuronas del núcleo subtalámico (NST) (Fig. 2). Así, el disparo de las neuronas espinosas de la red indirecta tiene como objeto desinhibir (aumentar la actividad) a las neuronas del NST. Las neuronas del NST también disparan de manera sostenida pero son excitatorias (glutamatérgicas), y sus blancos principales son las neuronas de los núcleos de salida: SNr y GPi (Alexander, 1995), adonde mandan conexiones divergentes y difusas (Mink, 1996). Por lo tanto, la desinhibición del NST producida por un comando cortical, aumenta la actividad de las neuronas de GPi/SNr. Esto produce una sobreinhibición de los circuitos tálamo-corticales que inhibe la ejecución de movimientos. Si el NST se lesiona, aparecen movimientos involuntarios repentinos e incontrolados (balismos).
El NST también recibe entradas directas de la Cx motora. Por lo tanto, este núcleo hace un balance entre las entradas facilitadoras de la Cx motora y entradas inhibidoras del GPe. Sólo cuando el NE "da su permiso", e inhibe a las neuronas del GPe, se facilitan las entradas corticales y se activa con más fuerza el NST; ésto redunda en una fuerte inhibición de circuitos tálamocorticales del movimiento para ciertos grupos musculares. Sin embargo, las entradas corticales al NE son de toda la Cx (sensorial y motora), mientras que las de Cx sobre el NST sólo son de la Cx motora. El NST, entonces, se encarga de equilibrar la actividad de diferentes poblaciones corticales: unas se inhiben focalmente, mientras otras mantienen el tono.
Resumiendo, la red directa activa los circuitos tálamo-corticales del movimiento por desinhibición, mientras la red indirecta inactiva los circuitos tálamo-corticales del movimiento por un aumento de la inhibición aportada por los núcleos de salida que son activados por el NST. Así, los núcleos de salida parecen equilibrar ambas redes, directa e indirecta, que se oponen a este nivel.
Papel de la dopamina en la red indirecta
Pero sucede que las neuronas espinosas de la red indirecta poseen receptores dopaminérgicos distintos a los que expresan las neuronas de la red directa. Las neuronas de la red indirecta expresan mayoritariamente receptores del tipo D2. Datos preliminares (no publicados) muestran que la activación de los receptores D2 reduce las corrientes persistentes de calcio de tipo L. Por lo tanto, su activación no favorece la excitación cortical, si no que la frena. Así, la DA facilita a las neuronas de la red directa que activan los movimientos y deprime el disparo de las neuronas de la red indirecta que inhiben los movimientos. En ambos casos, el resultado final en los núcleos de salida es el de favorecer la activación. El grado de "oposición" o equilibrio entre las redes directa e indirecta depende entonces de la DA. Ahora bien, los receptores D2 tienen mayor afinidad por la DA que los receptores D1 y están tónica y parcialmente activados por la liberación basal de DA (Schultz, 1998). En cambio, los receptores D1 se activan por elevaciones fásicas de la concentración de DA (Gonon, 1997; Hernández-López et al., 1997). Esto sugiere que la actividad tónica y generalizada sobre la actividad muscular es modulada por los receptores D2 (red indirecta), mientras que el efecto fásico de la DA sobre los receptores D1 (red directa) es el que selecciona las poblaciones de neuronas a activar (Mink, 1996). La red indirecta serviría entonces para "ajustar el borde" o frontera entre las neuronas activadas y las no activadas (como el botón de contraste de un televisor). Asimismo, la acción dual de la DA, la inhibición lateral favorecida por ella, y la activación focalizada de la Cx sobre el NST seleccionarían las distintas poblaciones neuronales que reducirían su activación o se inhibirían para permitir los movimientos (músculos antagonistas). Los mecanismos de control parecen estar a varios niveles.
La deficiencia de DA provocaría sobreactividad de la red indirecta e hipoactividad de la red directa. Si la actividad de la red indirecta fuera mucho mayor que la de su contraparte, habría una potente inhibición de los circuitos tálamocorticales del movimiento. Esto resultaría en acinesia, o dificultad para realizar movimientos. Este es otro signo de la enfermedad de Parkinson, y se alivia mediante la destrucción de las neuronas hiperactivas de los núcleos de salida en el GPi (palidotomía parcial).
En la enfermedad de Parkinson hay degeneración de las neuronas dopaminérgicas y depleción de la dopamina neoestriatal. Esto entorpece todo el funcionamiento de la red aunque las conexiones que utilizan neurotransmisión rápida (glutamato y GABA) estén intactas. Hay coactivación de grupos musculares pues no se separan bien las poblaciones neuronales de los músculos agonistas y antagonistas, lo que provoca rigidez. Hay sobre-excitación de la red indirecta respecto de la directa, lo que ocasiona acinesia; las motoneuronas que poseen conexión directa con las piramidales de Cx (<10%), las de los músculos distales de los dedos y las extremidades, se ven activadas cíclicamente sin regulación inhibitoria, lo que produce el "temblor en reposo" característico.
Epílogo
Nos hemos enfocado en las acciones de la DA sobre las neuronas de proyección del NE y sus entradas excitatorias e inhibitorias. Sin embargo, existen otros receptores a la DA en las neuronas neoestriatales tanto de proyección como interneuronas (D3 y D4). La DA interactúa con otros transmisores importantes tales como la acetilcolina, la substancia P, la somatostatina, las encefalinas, etc. Por último, la DA también actúa en otros núcleos de los GB y la Cx, e.g., STN, GPe, GPi, etc. Además, los GB no sólo proyectan al tálamo, también lo hacen al colículo superior y a la formación reticular (núcleos pedúnculo pontinos) influyendo directamente sobre vías descendentes a la ME (Fig. 2). Sin embargo, esta descripción parcial hace evidente lo difícil que es entender la transmisión a nivel central: un mismo transmisor modulador puede producir acciones tan complejas y diversas sobre tan diversos receptores, conductancias iónicas y neuronas de un circuito, que es capaz de reconfigurar toda la actividad del mismo. Por consiguiente, la falta de un sólo transmisor puede llevar a trastornos globales en el funcionamiento de toda la red, como es el caso de la enfermedad de Parkinson.
Referencias Bibliográficas
Aceves, J., Floran, B., Sierra, A., Mariscal, S. (1995). Receptor mediated modulation of the release of gamma-aminobutyric acid by endogenous dopamine in the basal ganglia of the rat. Prog-Neuropsychopharmacol-Biol-Psychiatry. 19: 727-39.
Alexander, G.E. (1987) Selective neuronal discharge in monkey putamen reflects intended direction of planned limb movements. Exp. Brain Res. 67: 623-634.
Alexander G.E. (1995) Basal Ganglia. En: The Handbook of Brain Theory and Neural Networks. (Arbib M.A., Ed). pp. 139-144. The MIT Press, Cambridge, MA.
Cepeda, C., Levine M.S. (1998) Dopamine and N-methyl-D-aspartate receptor interactions in the neostriatum. Dev. Neurosci., 20: 1-18.
Flores-Hernández, J., Galarraga, E., Bargas, J. (1997) Dopamine selects glutamatergic inputs to neostriatal neurons. Synapse. 25:185-195.
Galarraga, E., Hernández-López, S., Reyes, A., Barral, J., Bargas, J. (1997) Dopamine facilitates striatal EPSPs through an L-type conductance. NeuroReport 8: 2183-2186.
Gonon, F., (1997) Prolonged and extrasynaptic excitatory action of dopamine mediated by D1 receptors in the rat striatum in vivo. J. Neurosci.17: 5972-5978.
Hernández-López, S., Bargas, J., Surmeier, D.J., Reyes, A., Galarraga, E. (1997) D1 receptor activation enhances evoked discharge in neostriatal medium spiny neurons by modulating an L-type Ca2+ conductance. J. Neurosci., 17: 3334-3342.
Mink, JW (1996) The basal ganglia: focused selection and inhibition of competing motor programs. Prog. Neurobiol., 50: 381-425.
Pacheco-Cano MT, Bargas J, Hernández-López S, Tapia D, Galarraga E (1996) Inhibitory action of dopamine involves a subthreshold Cs+-sensitive conductance in neostriatal neurons. Exp. Brain Res., 110: 205-211.
Schultz, W. (1998) Predictive reward signal of dopamine neurons. J. Neurophysiol. 80: 1-27.
Surmeier DJ, Bargas J, Hemmings HC, Nairn AC, Greengard P (1995) Modulation of calcium currents by a D1 dopaminergic protein kinase/phosphatase cascade in rat neostriatal neurons. Neuron, 14: 385-397.
Suszkiw, J.B. (1996) Motor Funtions of the brain. En: Essentials of Physiology. 2nd. Ed. (Sperelakis, N., Banks, R.O. Ed.). pp: 141-152. Little Brown. Boston.
Wickens, J.R., Oorschot, D.E. (2000). Neural dynamics and surround inhibition in the neostriatum: a possible connection. En: Brain Dynamics and the Striatal Complex. (Miller, R. y Wickens, J.R. Ed.). pp: 141-149, Harwood Academic Publishers, Australia.
Wilson C.J. (1998) Basal Ganglia. En: The Synaptic Organization of the Brain (Shepherd G.M. Ed.). pp: 329-375, Oxford University Press, Oxford.
