BIOLOGIA |
Comparaciones en Fisiología y Evolución,
|
Visión de colores
El premio Nobel, Karl von Frisch (1886-1982) no sólo desentrañó el misterio de la comunicación en la danza de las abejas, sino también nos abrió una ventana al peculiar y diferente mundo visual de estos animales. Por ejemplo, descubrió que las abejas pueden ver luz polarizada. Y aún antes, cuando se suponía que sólo los humanos poseían visión de colores, von Frisch descubrió que ellas también veían colores. Un par de años después, su contemporáneo A. Kühn encontró que ellas podían ver luz ultravioleta. Igualmente importante fue en 1924 la fundación por von Frisch y Kühn de la revista Journal of Comparative Physiology, creando así un foro para un campo de estudio reciente en pleno desarrollo. A través de la comparación de las capacidades sensoriales de un vasto rango de animales, los fisiólogos comparativos deseaban no sólo descubrir algunas extrañas capacidades en sus sujetos, sino también generar un panorama más completo de los mecanismos sensoriales presentes en el reino animal. En estas dos direcciones, con cuidado e ingeniosos experimentos, los fisiólogos comparativos fueron extremadamente exitosos. Sin embargo, lo fueron menos al explicar el valor adaptativo de los mecanismos que encontraron, lo cual era sin duda una de las metas que se propusieron. En realidad, sus explicaciones del por qué los animales podían hacer las cosas que hacían, se limitaban en la mayoría de los casos (pero no siempre) a especulaciones de estilo barroco y pan-adaptacionista. Esto se debió a que algunos investigadores trataban cada especie como una entidad fuera del contexto evolutivo o histórico. En un sentido, se pensaba que cada especie se adaptaba individualmente a su ambiente particular, y que sus rasgos variaban libremente en todas las direcciones. Otros fisiólogos compararon abejas con humanos (u otras especies relacionadas) y las preguntas que surgían de estas comparaciones eran obvias. Por ejemplo, ¿por qué los insectos (y otros artrópodos) pueden ver luz ultravioleta y polarizada en tanto que nosotros no? La respuesta aparecía igualmente evidente: debe haber algo en el ambiente de los animales que hace que estos malabares sean ventajosos.
¿Pero es esto realmente así? Un ejemplo ilustrará este problema. En un estudio reciente, Brownell & Weber (1995) descubrieron que dos especies estrictamente nocturnas de escorpiones eran sensibles a luz polarizada. Este hecho intrigó a Brownell & Weber, dado que en la noche el cielo no contiene ningún índice de luz polarizada. Pero, debido a que seguramente había una razón adaptativa, ellos siguieron explorando la ecología visual de estas especies nocturnas para encontrar una explicación. Desafortunadamente, los receptores sensibles a luz polarizada no son una adaptación de los escorpiones, o de cualquier otro artrópodo. La sensibilidad a la polarización es simplemente una propiedad intrínseca en la construcción rabdomérica de los fotorreceptores. Dado que a menudo esta sensibilidad a la polarización no es deseable, algunos invertebrados han inventado algunos trucos para eliminarla (Wehner & Bernard 1993), los cuales son verdaderas adaptaciones; no lo son en cambio la sensibilidad a la polarización en un fotorreceptor de artrópodo. La respuesta a la pregunta de por qué los escorpiones nocturnos ven luz polarizada es, simplemente, porque sus ancestros lo hacían.
¿Podemos inferir la manera como las especies extinguidas veían el mundo? Puesto que los proyectos sobre las máquinas del tiempo habitualmente generan complicaciones, hoy en día es difícil obtener fondos para financiarlas. Sin embargo, los biólogos evolucionistas adoptaron una estrategia alternativa llamada el método comparativo (Harvey & Pagel 1991). Este análisis se basa en la observación de que los rasgos biológicos tienden más bien a conservarse que a cambiar. Si dos taxas hermanas comparten un rasgo común, es más probable que este rasgo se haya desarrollado en un ancestro común que de manera independiente en las dos especie. Sin embargo, dado que todos los artrópodos ven luz polarizada, los escorpiones nocturnos no desarrollaron particularmente esta capacidad. Ellos la tuvieron desde un comienzo y no hubo presión selectiva para deshacerse de ella. En un caso así no es pertinente buscar una explicación que implique adaptación. Este simple razonamiento evolutivo es el que está ausente en numerosos estudios fisiológicos. Inexplicablemente, algunos investigadores no han motivado a los fisiólogos a interesarse en los árboles filogenéticos: "la fisiología comparativa necesita más la colaboración de la ecología fisiológica que de la historia evolutiva o sistemática. En realidad.... la fisiología comparativa parece principalmente dependiente... de explicaciones adaptativas " (Waterman 1975). Como los ejemplos anteriormente señalados sugieren, esta posición es difícil de sostener. La fisiología comparativa necesita de la sistemática para entender la adaptación. Basándonos en dos ejemplos clásicos de von Frisch- la visión de colores y la danza de las abejas- apoyaremos esta posición.
El descubrimiento de que las abejas ven la luz ultravioleta (UV) y que las flores reflejan esas longitudes de onda corta fue hecha en los años 20 (ver referencias en von Frisch 1965). Desde entonces que los científicos han especulado que en abejas, los receptores al UV se desarrollaron a partir de un proceso coevolutivo con la coloración de la flora. Recientemente esta idea cobro un interés aún mayor, cuando modelos computacionales sugirieron que el sistema de visión de colores en abejas es el sistema óptimo para detectar e identificar flores (Chittka 1996a). ¿Pero es realmente recíproca la influencia evolutiva entre abejas y flores? Para probar que las señales de las flores dirigieron la evolución de la visión de colores en abejas, habría que mostrar que los ancestros de las abejas anteriores a las Angiospermas poseían diferentes tipos de receptores para el color. ¿Pero, cómo podemos determinar de qué color veían los insectos el mundo hace 200 millones de años? Para responder a esta pregunta uno tendría que estudiar miembros de la taxa de los artrópodos cuyos lineajes evolutivos divergieron del de las abejas con anterioridad a la aparición de las flores. Si la visión de colores de estos animales es indistinguible del de las abejas, implicaría que un tal sistema ya se encontraba presente en un ancestro que precedió a la evolución del color de las flores. Afortunadamente, los fisiólogos comparativos han reunido una gran cantidad de información sobre los receptores para el color en varias taxas de artrópodos (Menzel et al. 1986). Nosotros sólo necesitamos representarlos en un árbol filogenético, para que todo patrón de adaptación se haga de inmediato aparente.
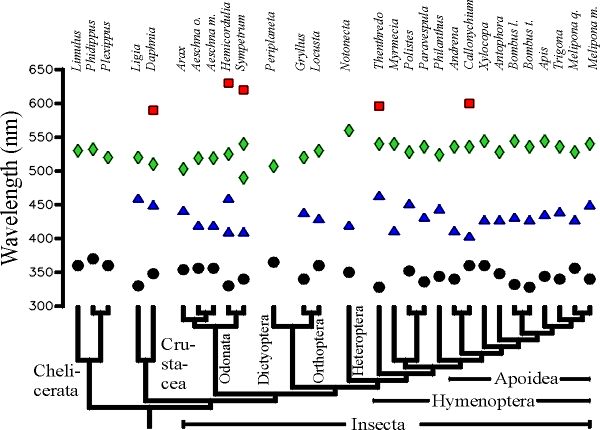
Fig. 1 Muestra los valores de lmax en fotorreceptores de 29 especies de artrópodos sobrepuestos al árbol filogenético de esas especies. O - receptores al UV (325-370nm); D - receptores al azul (400-460nm); à - receptores al verde (490-560nm); o - receptores al rojo (590-630). El nombre completo de las especies y una descripción de los resultados y citas originales se encuentra en Chittka (1996a).
Los valores lmax en Crustacea e Insecta se agrupan en tres distintos grupos alrededor de 350, 440 y 520 nm (Fig. 1). La mayoría de los Chelicerata no tienen receptores para el azul. En contraste, casi todos los Mandibulata (Crustacea e Insecta) poseen al menos tres tipos de receptores para el color con sensibilidad máxima en el UV, azul y verde (Chittka 1996b). Así, el receptor para el azul parece ser una novedad evolutiva en el ancestro de los Mandibulata. Las especies de insectos en las cuales uno de los tres tipos de receptores está ausente (Periplaneta y Myrmecia) representan casos claros en los cuales esos receptores fueron perdidos secundariamente. Los receptores al rojo aparecen de manera irregular en crustáceos e insectos, los cuales han aparecido varias veces de manera independiente durante la evolución. La posición lmax en los receptores UV, azul y verde se encuentran sorprendentemente conservados en Mandibulata. Así, podemos inferir que los ancestros del período Cámbrico de los insectos y crustáceos actuales poseían receptores para el UV, azul y verde, mientras que el ancestro común de los Mandibulata y Chelicerata no tenía receptor para el azul. Los insectos estaban preadaptados para codificar el color de las flores, hace más de 500 millones de años atrás, cerca de 400 millones de años antes que la extensiva radiación de las plantas con flores que comenzaron en medio del Cretáceo (hace 100 millones de años atrás), aunque los orígenes quizás tengan que situarse en el Triásico. Datos recientes sobre la estructura molecular de fotopigmentos apoyan la interpretación de que los tipos básicos de pigmentos visuales en artrópodos se encontrarían ya en las raíces de la evolución de los Artrópodos (A. Briscoe, comunicación personal). ¿Por qué la posición de lmax en los fotorreceptores es tan conservada en artrópodos? Los fotopigmentos con lmax en determinadas regiones del rango espectral entre 320 y 630 nm no constituye una imposibilidad técnica, ya que hay algunas especies de Crustácea e Insecta que poseen retinas con más de 5 tipos de pigmento visual, algunos de los cuales llenan los "espacios vacíos" en la escala de longitudes de onda comúnmente no ocupadas por los lmax de artrópodos. Una explicación más plausible es que la variabilidad genética para modificar el máximo de absorción del pigmento no estuvo presente en las especies en cuestión (Goldsmith 1990). En efecto, los estudios moleculares en pigmentos visuales en insectos muestran que mutaciones en diferentes posiciones son necesarias para modificar la sensibilidad espectral de un fotorreceptor (Britt et al. 1993).
En resumen, la respuesta a la pregunta de por qué las abejas poseen receptores al UV es la misma que arriba: porque sus ancestros los tenían. La vasta distribución de receptores al UV, azul y verde en artrópodos puede ser adaptativa, hacia una serie de parámetros ambientales no identificados. La hipótesis de que la visión de colores en insectos se adaptó a una clase particular de objetos, por ejemplo las flores, debe ser rechazada para el caso de las abejas. A lo más, la tricromacía UV, azul, verde puede constituir una adaptación general que permite al menos una codificación local óptima para una serie de objetos naturales y bajo diversas condiciones de iluminación (Chittka 1996a).
Las danzas de las abejas
El análisis del lenguaje de la danza en las abejas de von Frisch es todavía uno de los descubrimientos más fascinantes en comunicación no humana. Fuera de los humanos, las abejas aparecen como las únicas que se apoyan en un lenguaje simbólico para transmitir a sus congéneres la posición de una fuente de alimento provechosa. Después de que los forrajeros exitosos vuelven a la colmena, desarrollan una clara serie de patrones motores estereotipados en forma de ocho. La velocidad de esas danzas codifica la distancia y la dirección de la posición en relación a la gravedad, codifica la dirección de la comida en relación al sol. El lector encontrará en von Frisch (1965) una excelente y detallada monografía sobre "la danza de las abejas". ¿Como evolucionó esta fascinante conducta? Sorprendentemente, han habido intentos de entender la evolución de la danza de las abejas fuera de todo razonamiento filogenético (Gould & Towne 1986). Como se esperaba, estos intentos no han sido exitosos, al igual que los intentos de entender la evolución del lenguaje de las abejas en base a estudios de conductas similares en insectos solitarios así como las moscas o polillas (von Frisch 1965). Esas observaciones, aunque interesantes, no pueden ser usadas en un análisis evolutivo dado que la relación filogenética entre esas especies es demasiado lejana. El estudiante más exitoso de von Frisch, Martin Lindauer, estaba en lo cierto cuando planteó que la respuesta podría ser encontrada solamente si se comparaban especies cercanas filogenéticamente a nuestras abejas de la miel (ver von Frisch 1965). La reconstrucción de Lindauer de la evolución de la danza de las abejas es una obra maestra en cuanto al seguimiento de la evolución de rasgos de conducta compleja. El examinó tres especies tropicales de abejas de la miel y una variedad cercana a esta última, la abeja sin aguijón (stingless). Nosotros aludiremos aquí sólo brevemente a estos resultados porque ellos han sido descrito en otra parte (Lindauer & Kerr 1958, von Frisch 1965).
El panorama que emerge de los estudios de Lindauer es el siguiente: en todas las especies del género Apis las danzas consisten en patrones motores parecidos que contienen toda la información necesaria sobre la distancia y dirección de la fuente de comida. La dirección es expresada relativa a la posición actual del sol. En todas las especies, excepto en una considerada como la más "primitiva" (Apis florea), este ángulo es expresado durante la danza (la cual es naturalmente efectuada en una superficie vertical) como el ángulo relativo a la gravedad. Apis florea, especie que anida a campo abierto y cuyos miembros danzan en una superficie horizontal, no realizan la transformación relativa a la gravedad, sino más bien sus miembros orientan sus danzas en relación a la dirección del sol, cuando emprenden el vuelo hacia el comedero. Lindauer pensó que ésta era la forma original de la danza. Lo cual no es completamente verdadero. Una premisa entre los biólogos no evolucionistas es que las especies existentes y que divergieron tempranamente a partir de un tronco común (especies más "primitivas") deberían conservar los patrones ancestrales de conducta u otros rasgos y así, en el caso de las abejas, dar cuenta de una manera directa de las formas ancestrales de la danza que von Frisch observó en la abeja Apis mellifera. Pero las especies ancestrales han tenido tanto tiempo desde el punto de divergencia como las especies derivadas (que emergen de las especies hermanas de las ancestrales) para desarrollar un sistema de comunicación y, al igual que en las especies cuya morfología y hábitos de nidificación se han mantenido constantes desde el tiempo de divergencia, la conducta puede haber adoptado cambios adaptativos. La reconstrucción de la danza ancestral es posible sólo a través de la identificación de caracteres compartidos entre taxas hermanas, y la identificación de grupos externos que carecen de esos rasgos. En una reciente reconstrucción filogenética de la evolución de la danza en el género Apis, Dyes (1991) señala que es equivalente asumir que A. florea perdió secundariamente la habilidad para la transformación.
¿Nos pueden dar las abejas sin aguijón (grupo hermano de las abejas de la miel), información acerca de cual escenario es el correcto? Desafortunadamente no. Ninguna de las numerosas especies parece realizar el tipo de patrón motor repetitivo llamado danzas en la abeja de la miel. La mayoría, sin embargo, realizan carreras con cierto grado de excitación en el panal al regresar de un forrajeo exitoso (Lindauer & Kerr 1958). Algunas especies también emiten pequeñas pulsaciones vibracionales durante esas carreras, las cuales están correlacionadas con la distancia a una fuente de comida, al igual que en las abejas de la miel (Esch 1967). Con ellas también comparten el uso de la trofilaxis: a menudo los forrajeros exitosos regurgitan el néctar durante sus danzas (o carreras excitadas, si Ud. prefiere no llamar danza a este caótico patrón motor en las abejas sin aguijón). Al lado de estos rasgos comunes, las abejas sin aguijón muestran marcadas diferencias en el sistema de reclutamiento entre especies. Algunas usan huellas aromáticas, y algunas guían a sus compañeros de nido directamente a la fuente de alimento. En conclusión, el ancestro común de las abejas sin aguijón y de las abejas de la miel aparentemente ejecutaban carreras excitadas en el panal luego de encontrar el alimento, mostraban trofilaxis, y usaban sonidos para alertar a los compañeros de nido y posiblemente codificaban la distancia. La manera de como el ancestro de la abeja de la miel desarrolló una forma más elaborada de la danza - cuyo descubrimiento llevó a von Frisch a ganar el premio Nobel- quizás jamás será conocida. Hay simplemente una distancia demasiado grande entre la conducta de la abeja de la miel y la de sus parientes más cercanos. Algunos investigadores han sugerido que la codificación de la dirección en la danza de la abeja de la miel puede haber evolucionado a partir de "movimientos planificados" presentes en algunas especies de abejas sin aguijón (von Frisch 1965, Esch 1967). La abejas sin aguijón del género Melipona repiten "falsas partidas" en la dirección del alimento, pero se devuelven sólo para despegar otra vez. Pese a que Melipona es considerada un "género avanzado" de abeja sin aguijón, no es el pariente más cercano de las abejas de miel. En cambio, el pariente más cercano de los ancestros de la abeja de la miel es el ancestro de todas las abejas sin aguijón. Dado que las falsas partidas no se encuentran en todas las abeja sin aguijón, seguramente estas no corresponden a la condición más ancestral, no podemos afirmar que estas sean ancestrales en el grupo de hermanas de las abejas sin aguijón y las abejas de la miel. Así el origen de la danzas en la abeja de la miel, incluyendo su codificación simbólica de la distancia y dirección, quedará sin duda como un misterio.
El verdadero origen de las "danzas" en los ancestros del Cretáceo de las abejas eusociales Apid (abeja de la miel, abeja sin aguijón y abejorro) está aún en penumbras. ¿Cómo establecieron por primera vez el contacto los danzadores y los reclutas? ¿Cuál evolucionó primero la peculiar conducta de los forrajeros exitosos de correr excitados alrededor del panal, o la conducta de alerta de los reclutas para seguir a los forrajeros exitosos? Claramente, las dos necesitan existir para que el reclutamiento funcione. ¿Pero por qué una abeja cualquiera seguiría a un cazador exitoso si tal cazador aun no ha "inventado" un mensaje que al final lo identifique como tal? Y seguramente, este es, aunque difícil de concebir, equivalente a que el cazador exitoso desarrolle patrones motores adecuados sin que nadie este presente en el panal (dado que las abejas del tiempo de la pre-danza no habían aun "inventado" su danza con conducta de seguimiento). von Frisch (1965) cita evidencias que el abejorro, el pariente más cercano de las abejas sin aguijón y las abejas de la miel, hacen exactamente lo mismo. Trabajos anteriores observaron que los abejorros al regreso de un comedero darían vueltas de una manera excitada similar a la observada en el panal, y ocasionalmente chocan a los compañeros de panal (como la mayoría de las especies de abeja sin aguijón lo harían) - pero a nadie le importan esas maniobras - otras abejas no parecen impresionadas por tales forrajeros excitados y podrían no ser reclutadas hacia la fuente de alimento. Examinamos tres especies de abejorro (Bombus impatiens, B. occidentalis, y B. terrestris) y observamos conductas similares a las previamente descritas: una abeja exitosa que regresa no sólo descarga su captura, sino que realiza también largas carreras a través del panal, arriba y abajo de las paredes del nido, y algunas veces pueden también dejar el nido sólo para regresar y realizar más idas y vueltas al panal. Esta conducta puede durar algunos minutos antes que el cazador encabece el retorno al comedero. Pero ninguna de las abejas en el nido sigue a la abeja que "baila". ¿Podría esto entonces significar que "danzar" realmente precede la evolución de la danza de seguimiento? ¿Empezaron los danzadores enviando mensajes antes de que alguien los "escuche"? Si la adecuación de la colonia depende de la ingestión de comida, ¿por qué los forrajeros perderían su precioso tiempo el cual podría ser gastado colectando comida? Si no hubiera una ventaja de adecuación inmediata para tal extraña conducta, ¿cómo podría ésta haber evolucionado?
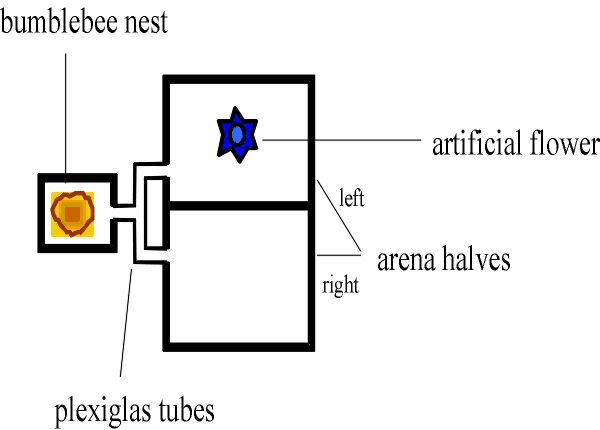
Fig. 2. Dispositivo para medir si el abejorro muestra conducta de reclutamiento. Una abeja es transportada de manera continua entre el nido y un comedero en la mitad izquierda del campo de ensayos, para recolectar tanto néctar como pueda. Para verificar si ella transmitía información en el nido sobre la presencia de los comederos (más que a nivel del comedero) nosotros desviamos todos los reclutas hacia el lado derecho de la arena (bumblebee nest = nido del abejorro; plexigalss tube = tubo plástico; left = izquierda; right = derecha; arena halves = mitades del campo de ensayos; artificial flower = flor artificial).
Sospechamos que, después de todo debe haber un mensaje. Para evaluar esta posibilidad, se dejó que un abejorro colectara néctar en un comedero localizado en una arena de vuelo (un dispositivo experimental de 40cm x 60cm con una cubierta transparente para la observación de la conducta de la abeja). Una vez que la abeja descubría el néctar, ella viajaba continuamente entre el comedero y el panal para aportar una mayor cantidad de cargas de alimento. Durante su estadía en el panal, observamos las interacciones con sus compañeras de nido (Dornhaus et al. 1999). Encontramos que, pese a que otras abejas no prestaban atención a las "danzas" de la abeja, sí estaban muy interesadas en la comida que ella traía al panal. Inmediatamente después que ella descargaba su carga en uno de las numerosos celdas de miel en el nido, otras abejas examinaban el nuevo néctar. ¿Como sabían que el nuevo néctar había sido traído? De alguna manera el cazador a su regreso debe haber avisado a las otras abejas. Pero aun más sorprendentemente, encontramos que si la abeja experimental traía varias cargas de forraje a casa, la colonia completa de forrajeros salía del nido en busca del alimento. Explícitamente, nos aseguramos que la abeja recompensada no pudiera reclutar otras abejas marcando de olor el comedero. Esto lo hicimos desviando las abejas reclutadas hacia una arena diferente. Así las danzas primitivas no fueron sin sentido - estas sirvieron para motivar otras abejas a buscar alimento. ¿Pero, cómo fue transmitido el mensaje, si no a través de un contacto mecánico? Una posibilidad puede haber sido que las abejas que esperaban en el panal simplemente reaccionaron a un aumento del abastecimiento de néctar, y tomaron esto como una indicación que fuentes ricas en alimentos habían sido descubiertas. En efecto, fuimos capaces de imitar el efecto de un regreso exitoso de forrajeo regular introduciendo un "forrajero fantasma": regularmente se introdujo pequeñas cantidades de néctar en los lugares de miel de la colonia por medio de una jeringa, elegimos las cantidades de manera que éstas se ajustaran a las que aporta una abeja cazadora normal. La abeja fantasma motivó a varias abejas - pero no tantas como en el caso de una abeja real - a abandonar el nido y buscar. Debe haber habido una señal adicional transmitida mientras que la abeja recompensada daba vueltas alrededor del panal. Para verificar que señal podría haber sido, pusimos dos nidos uno al lado del otro, pero separados por medio de dos capas de malla de alambre, alejadas suficiente para impedir que las abejas de las dos colonias tuvieran contacto entre ellas. Luego, recompensamos las abejas de un panal ofreciéndoles alimento en la arena de vuelo, y contamos el número de abejas entrando en la otra arena conectada al segundo panal. En efecto, las abejas en el panal no recompensado dejaron su casa en búsqueda de alimento cuando la colonia vecina fue recompensada. Estas observaciones sugieren que la abeja exitosa emite una feromona que informa a otras abejas de la presencia de interesantes fuentes de alimento. También es posible que las abejas emitan vibraciones en el substrato que alerten otras cazadoras, como es el caso en las abejas de la miel y sin aguijón. En realidad, los abejorros forrajeros emiten breves sonidos cuando regresan al panal, pero esos sonidos son independientes de si el regreso fue exitoso e independiente de lo lejos que la abeja tuvo que viajar. Los sonidos son producidos también cuando la abeja abandona brevemente el panal, sin siquiera volar hacia la fuente de la comida (Heidelbach et al. 1998, H. Böhm, comunicación personal). Cuando separamos los dos panales con una sabana de plástico, manteniendo así contacto visual entre las dos colonias y la posibilidad de transmisión por vibración, encontramos que el panal sin recompensa dejaba de responder. Entonces, las carreras excitadas de los abejorros forrajeros (posiblemente el ancestro de todas las abejas eusocial) sirve aparentemente al propósito de distribuir de una manera tan general como posible un "mensaje" de feromona hacia toda la colonia. Admitiendo que este escenario es especulativo, por el momento no existe otra mejor interpretación para explicar por qué los forrajeros que han descubierto fuentes de alimento deben invertir su tiempo en ejecutar patrones motores articulados si no hay la recompensa de la atención al respecto. Obviamente, nuestra explicación podría ganar más peso si pudiéramos mostrar que tales feromonas son emitidas durante la danza en otras abejas sociales, así como en la abejas sin aguijón y en las abejas de la miel. Existe una evidencia preliminar que muestra que en la abeja de la miel este es el caso (J. Tautz, comunicación personal). Claramente, necesitamos también datos en abejas sin aguijón. Una vez que dispongamos de estos datos, confiamos que pronto podría ser posible reconstruir los verdaderos orígenes de la comunicación en abejas.
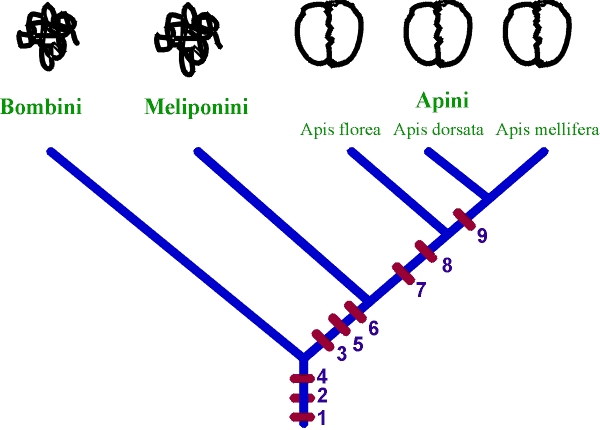
Fig. 3 Eventos mayores en la evolución de las danzas de las abejas, representadas en el árbol filogenético de las abejas eusociales. Las estructura básica de las "danzas" en varias taxa se muestran arriba. 1. carreras aceleradas en el panal por forrajeadores exitosos; 2. Sonda de néctar para trabajadores que han sido traídos al panal; 3. Pulsos de vibraciones de sonido producida por los forrajeadores que regresan; 4. Trofilaxis; 5. Largo de los zumbidos correlacionados con la distancia del alimento.; 6. "Danza" de seguimiento, o al menos "respuestas de rotación" hacia el cazador exitoso; 7. Patrón de movimiento con figura en ocho; 8. Danza ejecutada en una superficie horizontal; 9. Movimiento de danza ejecutada en panales verticales. Un evento adicional ocurrido a la raíz del árbol puede haber sido la expansión de una feromona distribuida por los forrajeadores exitosos.
Esperamos haber convencido el lector de que las comparaciones entre taxas hermanas son necesarias para trazar la evolución de la comunicación en abejas, y para esto, obviamente necesitamos la sistemática. Claramente, nuestras interpretaciones dependen entonces del árbol filogenético en particular que usemos como base. Si resulta que los abejorros, y no las abejas sin aguijón, son los parientes cercanos de las abejas de la miel, esto podría cambiar considerablemente nuestro entendimiento del sistema de reclutamiento en las abejas. Pero en este momento, aparece que la filogenia es más bien como la mencionamos anteriormente (Roig-Alsina & Michener 1993). Afortunadamente, se ha hecho también claro que nosotros no podemos entender la adaptación mirando a las especies como entidades aisladas que puedan libremente variar sus rasgos de conducta para adaptarse a cualquier condición ambiental. Por ejemplo, el lenguaje de la danza de la abeja de la miel Apis mellifera ha sido considerado particularmente eficiente para explotar el mercado de las flores de hábitats temperados (von Frisch 1965). Aunque esto debería ser verdadero (lo cual es difícil de medir), esto claramente no significa que el lenguaje de la danza evoluciona en Apis mellifera como una adaptación para la comunicación floral en un clima temperado. Es muy probable que el ancestro de todas las abejas de la miel vivía en los trópicos, y que el lenguaje evolucionó en ese ancestro (Dyer 1991). Entonces ésta no es una adaptación a algo temperado (aunque es improbable que esto sea una desventaja en tales hábitats). ¿Pero es el lenguaje de la danza una adaptación a un sector particular de fuente alimenticia floral en hábitats tropicales, en el cual las fuentes de néctar arbóreo son más frecuentes? En tales condiciones el reclutamiento puede ser rápidamente localizado en un retoño de árbol florecido, antes que éste sea descubierto y devastado por otros forrajeadores. Desgraciadamente, las abejas sin aguijón viven también en selvas tropicales, y ellas no usan el mismo lenguaje. Algunas especies, pueden no haber "inventado" aun este lenguaje, otras posiblemente quizás nunca lo harán. Algunas especies de abejas sin aguijón son más eficientes en reclutar parejas del panal usando huellas aromáticas y vigilar a las reclutas escoltas, que las abejas de la miel usando sus danzas más elaboradas (Lindauer & Kerr 1958). En tales especies, un mutante que "introduce" las danzas de la abeja de la miel puede seleccionar en el sentido opuesto, porque puede transformar el forrajeo social menos eficiente (sin que el costo de la huella aromática tenga mayor sobrepeso que el costo de la danza).
En conclusión, las danzas de las abejas de la miel que conocemos claramente no son el único resultado posible de una adaptación evolutiva de la comunicación en abeja en hábitats tropicales. La moraleja a considerar es que a menudo las diferentes especies de animales viven en condiciones que difieren en varias dimensiones. A menudo también difieren en algunos rasgos, que pueden ser morfológicos, de conducta o fisiológicos. Para todo rasgo particular, en cualquier animal es generalmente muy fácil encontrar una condición ambiental única para la cual el rasgo podría haberse adaptado. Sin tomar las precauciones necesarias, las interpretaciones de tales correlaciones están condenadas a ser especulativas y claramente necesitamos tomar en cuenta la historia de las especies para entender su adaptación.
Finalmente, otra razón del por qué conocemos tan poco acerca de como algunos rasgos fisiológicos o de conducta evolucionan es la negligencia de algunos fisiólogos en interpretar la variación interindividual. A menudo esta variabilidad es tratada como ruido, que necesita ser eliminada promediando un gran numero de medidas provenientes de diferentes animales. Y animales que no despliegan la conducta supuesta algunas veces ni siquiera entran en el análisis de los datos. Cuando discutimos este problema con Lindauer, el nos contó la siguiente anécdota. En 1952, el trabajó con von Frisch preguntándose si las abejas de la miel pueden comunicar altitud en sus danzas. Con este propósito, entrenaron las abejas de una colmena situada en la parte baja de un valle a un comedero situado en un puente de 76m de alto. Von Frisch (monitoreando las danzas de la colmena) y Lindauer (quien vigilaba el comedero en el puente) se comunicaban por medio de un receptáculo colgando de una cuerda que contenía breves notas en papel. En una ocasión, von Frisch pidió, a través de una nota, la "EJECUCIÓN DE LOS SIGUIENTES MALOS DANZADORES" seguido de una lista de abejas tontas con la infortunada etiqueta. Esto es más que una anécdota acerca del análisis de datos en los años 50. Por varias décadas, los científicos han acumulado resultados a partir de muchos animales, y eliminado las respuestas que muestran desviaciones importantes de lo esperado. Algunas veces esto puede ser legítimo. Datos de conducta y de fisiología son a menudo tan ruidoso que extraer cualquiera información de estos no es posible sin promediar. Y desviaciones marcadas de las observación esperadas podrían a menudo significar que la medida, por ejemplo en electrofisiología, es imperfecta. A través de estos promedios hemos quizás perdido mucha información valiosa. Volviendo al ejemplo de arriba, ¿Puede ser que la razón principal del alto nivel de conservación en los receptores para el color en artrópodos exista simplemente porque no hay variabilidad entre individuos en la especie en cuestión? Un número importante de científicos han trabajo en, por ejemplo, los receptores para el color en las abejas de la miel, y los resultados difieren tanto dentro de como entre estudios. Pero el debate acerca de esas diferencias en su mayoría se focalizan en las posibles contribuciones de artefactos o de diferentes métodos fisiológicos (Menzel et al. 1986). De hecho, las dos contribuciones pueden añadir ruido a las medidas, pero desafortunadamente, la posibilidad de que la variabilidad interindividual pueda también contribuir no ha sido siquiera considerada. Por lo tanto, no sabemos siquiera si el sistema de visión de colores en abejas contiene el material bruto para su evolución.
J. Gould escribió acerca de von Frisch: "Sus trabajos pioneros inspiraron el descubrimiento en diversos animales de sistemas sensoriales de otra manera inimaginables: detectores infrarrojos en las culebras cazadoras nocturnas, sonares ultrasónicos en delfines y murciélagos, audición infrasónica en pájaros, y sensibilidad al campo magnético en una variedad de animales. Sin duda, otros sistemas quedan aún por descubrir. La lección es melancólica: somos ciegos a nuestra propia ceguera, y debiéramos tratar de no proyectar nuestras propias incapacidades hacia el resto del reino animal" (Gould 1980). No podríamos más que estar de acuerdo con esto. Si, además, tratamos de no cerrar los ojos a los recientes (y no tan recientes) desarrollos en biología evolutiva, pronto podríamos ser capaces de explicar por qué somos ciegos a algunas cosas que otros animales perciben. Por qué algunos animales pueden ver cosas a las cuales nosotros somos ciegos, y por qué el resto del reino animal carece de algunas de las capacidades que nosotros presumimos ser nuestras.
Referencias
Bibliografía
Britt, S. G., Feiler, R., Kirschfeld, K. & Zuker, C. S. (1993). Spectral tuning of Rhodopsin and Metarhodopsin in vivo. Neuron, 11: 29-39.
Brownell, P. & Weber, T. (1995). Polarization vision in nocturnal sand scorpions. In: Nervous Systems and Behavior. Eds: Burrows, M., Matheson, T., Newland, P.L. & Schuppe, H. Thieme Verlag, Stuttgart, p. 281.
Chittka, L. (1996a). Optimal sets of colour receptors and opponent processes for coding of natural objects in insect vision. J. Theor. Biol., 181: 179-196.
Chittka, L. (1996b). Does bee color vision predate the evolution of flower color? Naturwissenschaft, 83: 136-138.
Dornhaus, A., Hartmann, F. & Chittka, L. (1999). A window into the past: what bumble bees tell us about Cretaceous dances. In: Goettingen Neurobiology Report 1999, Eds: Elsner N. & Eysel U., in press.
Dyer, F. C. (1991). Comparative studies of dance communication: analysis of phylogeny and function. In: Diversity in the genus Apis. Ed: Smith, D.R., Westview Press, Boulder, pp. 177-198.
Esch, H. (1967). The evolution of the bee language. Scientific American, 216: 96-104
Frisch, K. v. (1965). Tanzsprache und Orientierung der Bienen. Heidelberg: Springer.
Goldsmith, T.H. (1990). Optimization, constraint, and history in the evolution of eyes. Quart. Rev. Biol., 65: 281-322.
Gould, J.L. (1980). Navigation by honey bees. In: Genes, Cells, and Behavior. Ed: N. Horowitz & E. Hutchings, W.H. Freeman, San Francisco, pp. 135-141.
Gould, J. L. & Towne, W. F. (1986). Evolution of the dance language. American Naturalist, 130: 317-338.
Harvey, P.H. & Pagel, M.D. (1991). The comparative method in evolutionary biology. Oxford University Press
Heidelbach, J., Böhm, H. & Kirchner, W.H. (1998). Sound and vibrational signals in a bumblebee colony (Bombus terrestris). Zoology: 101: 82.
Lindauer, M. & Kerr, W. (1958). Die gegenseitige Verständigung bei den stachellosen Bienen. Z. Vergl. Physiol., 41: 405-434.
Menzel, R., Ventura, D. F., Hertel, H., de Souza, J. M. & Greggers, U. (1986). Spectral sensitivity of photoreceptors in insect compound eyes: comparison of species and methods. J. Comp. Physiol. A, 158: 165-177.
Roig-Alsina, A. & Michener, C. D. (1993). Studies of the phylogeny and classification of Long-tongued bees (Hymenoptera: Apoidea). Univ. Kans. Sci. Bull. 55: 124-162.
Waterman, T.H. (1975). Expectation and Achievement in Comparative Physiology. J. Exp. Zool. 194: 309-344.
Wehner, R.& Bernard, G. D. (1993). Photoreceptor twist: A solution to the false-color problem. Proc. Nat. Acad. Sci. (USA), 90: 4132-4135.
Punteros de Interés
1. Película esquemática de la danza de las abejas
http://ourworld.compuserve.com/homepages/Beekeeping/beedance.htm
2. La danza de la abeja descifrada
http://www.bhs.mq.edu.au/psy/105/lectures/chris2/Week13_p1.html
3. Como las abejas ven el mundo
http://cvs.anu.edu.au/andy/beye/beyehome.html
4. Como las abejas ven las flores
http://gears.tucson.ars.ag.gov/ic/vision/bee-vision.html
5. Información acerca de los abejorros
