
BIOLOGIA  |
Mejoramiento Genético de Animales |
|
|
©Hugo
Montaldo y Nelson Barría,1998
|
El concepto de mejoramiento genético de animales (MGA) posiblemente
sugiere distintas imágenes en diferentes personas. En el plano práctico,
surge la idea de usar y combinar mejores razas y animales en las diversas
especies de animales domésticos, sin preguntarnos mucho acerca de
definir o evaluar el mérito o de cómo definir mejores. En
el plano científico, las ideas que aparecen con más frecuencia
están relacionadas con los últimos avances publicitados en
tecnología reproductiva y molecular, como la clonación (producción
de animales genéticamente idénticos) y otras manipulaciones
recientes de la reproducción y el uso de marcadores genéticos
del ADN (depositario de la información genética de los organismos)
para la selección. En realidad, la situación es algo diferente.
Este trabajo pretende dar una visión general de la disciplina
y describir algunos de sus métodos y sus problemas actuales en 1998.
Un intento de definición
El MGA consiste en aplicar principios biológicos, económicos y matemáticos, con el fin de encontrar estrategias óptimas para aprovechar la variación genética existente en una especie de animales en particular para maximizar su mérito. Esto involucra tanto la variación genética entre los individuos de una raza, como la variación entre razas y cruzas.
El MGA involucra procesos de evaluación genética y difusión del material genético seleccionado, en los cuales se pueden usar tecnologías reproductivas artificiales tales como la inseminación artificial (AI), la ovulación múltiple y transferencia embrionaria (OMTE), la fertilización in vitro de embriones, así como el uso de marcadores de ADN. Los dos principales problemas que nos formulamos en un programa de MGA son:
1.-Cómo definir el mérito (objetivo del programa) y,
2.-Cómo lograr este objetivo (sistema de evaluación, uso y difusión de la mejora a la población comercial).
Las dos herramientas primordiales del MGA son la selección (determinar cuáles individuos van a dejar descendencia) y los sistemas de apareamiento (determinar cómo los individuos seleccionados serán apareados).
La medición de la producción animal
La herramienta que más ha impactado el mejoramiento animal en el mundo es el control de producción. En efecto, la medición objetiva de la producción de los animales sirve para hacer evaluaciones de los mismos para la selección, evaluar las razas y cruzas, estimar los parámetros requeridos para los programas, medir aspectos económicos y optimizar el proceso.
En un principio, los sistemas de control fueron generalmente simples e incluyeron solo evaluaciones de cantidad (producción de leche, peso del vellón en ovejas, pesos corporales a determinadas edades en animales productores de carne). Más recientemente, las necesidades del mercado y el deseo de los criadores de tomar decisiones en base a un conjunto de características mas estrechamente relacionadas con el valor económico de la producción y la consideración de aspectos sociales en los objetivos de producción, han incorporado una serie diversa de características de calidad, como el contenido de grasa de la carne y proteína de la leche, características de la carne como el área de músculo en diversos cortes, la dureza, el porcentaje de grasa etc. Algunas de estas características son difíciles o imposibles de medir en el animal vivo, por lo que son evaluadas a través de métodos indirectos. Un ejemplo es el uso del scanner para medir características de composición corporal como el porcentaje de grasa en el animal in vivo. Otras características que se evalúan en los programas de mejoramiento son las relacionadas con la fertilidad, la supervivencia y la resistencia a enfermedades. En etapas más recientes se ha intentado medir variables asociadas con los costos de producción, como el costo de los alimentos y medicamentos. En general, los avances técnicos están permitiendo realizar mediciones más precisas y frecuentes e incluir características más estrechamente relacionadas con los objetivos de los programas genéticos.
El control de producción está generalmente asociado, salvo en el caso de sistemas de producción extensivos en rumiantes, a un completo y riguroso control genealógico, que implica el registro de los padres de cada animal en la población y permite estructurar el pedigrí necesario para el cálculo de los coeficientes de parentesco utilizados en las evaluaciones genéticas y en el cálculo de coeficientes de consanguinidad.
Un aspecto fundamental de estos sistemas de evaluación es el control de calidad; los errores, tanto de datos de producción como de pedigrí pueden afectar negativamente las tasas de mejoramiento genético. Por este motivo, en los países con mayor desarrollo en este campo, la participación de organismos oficiales (Universidades, organismos estatales), han sido claves para propiciar la aceptación de la información de campo, como veraz y válida. Este desarrollo históricamente ha estado ligado a un alto grado de participación del gobierno y las organizaciones de ganaderos. Esta naturaleza colectiva se ha requerido debido a la naturaleza a largo plazo de las inversiones en mejoramiento genético, a la necesidad de contar con personal capacitado para organizar y mantener en operación los sistemas de evaluación y a la necesidad en muchos casos de realizar las evaluaciones en los rebaños comerciales, como en el caso de los rumiantes.
En muchos países, intereses particulares de asociaciones de razas, agrupaciones gremiales, desconocimiento de los principios científicos de la mejora así como de una ineficiente coordinación y promoción de la mejora genética por parte del gobierno, han paralizado la aplicación de la mejora genética. El costo ecológico por el mantenimiento de grandes números de animales improductivos con un impacto negativo en el medio ambiente y en la economía para estos países, por la necesidad de importar material genético y por las pérdidas productivas, es difícil de cuantificar con precisión pero es sin duda considerable.
Modelos lineales + computadoras + inseminación artificial
El avance explosivo de la tecnología computación ligado a desarrollos teóricos en el área de los modelos estadísticos aplicados al análisis de datos obtenidos en condiciones reales de producción, permitieron la implementación de métodos muy eficaces para el cálculo de valores genéticos estimados (VGEs) aprovechando la información del control de producción y de pedigrí, en forma que actualmente se puede considerar que dichas implementaciones son una tecnología madura y su instrumentación es básicamente un ejercicio de aplicación.
Esta metodología ha permitido tremendos avances en algunas especies, como es el caso de los bovinos productores de leche. Se estima que una vaca promedio, obtenida por IA en Estados Unidos, incrementa su producción en más de 100 kg de leche por año (aproximadamente 0.32 lt/día) por razones puramente genéticas. Esto tiene, junto con el cambio en otras características, una gran importancia económica para los productores y la industria, al grado que los productos de las industrias que han realizado mayor mejora genética, como la producción de leche, huevos, carne de cerdo y carne de pollo, han capturado una mayor proporción del mercado, debido en parte, al abaratamiento de los costos de producción por la mayor eficiencia de los animales. Esto se ha logrado con el uso de animales superiores a través del uso de los VGEs para la selección. Estos VGEs son calculados con una metodología que se conoce como BLUP (best linear unbiased prediction o mejor predicción lineal insesgada), de acuerdo a las propiedades teóricas de las soluciones. Esto ha sido posible también gracias al desarrollo de métodos emparentados al BLUP como el REML (máxima verosimilitud restringida) para estimar parámetros genéticos o matrices de varianzas y covarianzas requeridas para ponderar adecuadamente los factores del modelo.
Dos descubrimientos claves en este sentido fueron las ecuaciones del modelo mixto de Henderson que redujo considerablemente las dimensiones de los sistemas de ecuaciones simultáneas a resolver para obtener los VGEs y la incorporación de las correlaciones entre los valores genéticos de los animales al modelo, mediante la matriz de parentesco, que se construye a partir de la información contenida en el pedigrí. Al obtener estimaciones confiables e insesgadas de los VGEs de animales en muchos rebaños, lo que fue posible al usar la inseminación artificial, por primera vez la industria pudo comparar los valores genéticos de los animales de toda la población, lo que le dio a la selección un tremendo poder. Este proceso tiende a destruir la estructura piramidal jerárquica tradicional de los hatos reproductores, debido a que los animales superiores son elegidos en forma horizontal.
En otros casos, la estructura jerárquica es mantenida parcialmente por una falta de adopción de las técnicas modernas como por ejemplo en la producción de lana en Australia o muchos productores de sementales de ganado de carne en América Latina. En especies con elevada fecundidad y reducido costo por individuo, el elevado número de progenie por animal reproductor y los menores costos de operación, permiten la centralización del proceso de mejoramiento en núcleos de empresas privadas que controlan todas las poblaciones de reproductores como en pollos de engorde, gallinas productoras de huevos y en forma creciente, cerdos. Una población rigurosamente seleccionada abastece en este caso a un estrato multiplicador, capaz de proveer de animales al estrato comercial. En cierto modo, una situación similar puede ser creada en rumiantes (a mayor costo), al incrementar su capacidad de reproducción usando la transferencia embrionaria y la fertilización in vitro.
Divide y vencerás, simplifiquemos el modelo
En cierto modo, el éxito de los métodos de selección es un producto de la simplificación. La aplicación de un modelo genético conceptualmente completo a características controladas por muchos pares de genes, implica considerar simultáneamente tantos parámetros correspondientes a efectos que complicarían el modelo en forma excesiva para hacerlo práctico. Los experimentos necesarios para generar este tipo de parámetros son frecuentemente imposibles o limitados a una escala que comprometería la utilidad de los estimadores. En un principio la simplicidad de los modelos de evaluación genética fue necesaria debido a limitaciones de capacidad de cómputo y la ausencia de una teoría matemática para realizar una estimación insesgada del valor genético en presencia de factores de confusión debidos al ambiente (años, épocas, rebaños) e información de parientes.
Los modelos de evaluación genética utilizan generalmente los efectos genéticos aditivos directos (del propio animal) y maternos (debidos a la variación del genotipo de la madre que afecta características que se miden temprano en la vida de los animales). Los efectos aditivos son aquellos que se deben al efecto independiente de cada alelo, mientras que la dominancia se debe a interacciones dentro cada locus y la epistasis a interacciones entre alelos localizados en diferentes loci. En términos generales la variación aditiva es la mas importante para la selección animal. Los efectos no aditivos son importantes en la manifestación de los efectos negativos de la consanguinidad y en los positivos de la heterosis. Los modelos utilizados actualmente para la evaluación dentro de una población contienen efectos genéticos aditivos y en ocasiones de dominancia. Esto no quiere decir que los efectos de interacción no sean importantes, solamente indica que en promedio son menos importantes bajo los modelos y suposiciones utilizados. El modelo puede ser una limitante, en el sentido de que sólo se puede encontrar lo que se busca.
En caracteres afectados por la variación genética maternal, como el peso al destete en rumiantes, generalmente los efectos maternos son incluidos en el modelo. En presencia de varias características, se usan enfoques multivariados que son extensiones de los modelos para una característica. Aquí un requisito crítico es la disponibilidad de estimadores adecuados de las correlaciones genéticas entre las características, que frecuentemente son difíciles de estimar con precisión.
Otra complicación importante son las interacciones genotipo
x ambiente, cuando los efectos genéticos y ambientales actúan
en forma conjunta para producir el fenotipo. Este tipo de interacción
puede reducir la eficacia de la selección realizada en otras condiciones
ambientales, debido a problemas de adaptación de los animales. Esto
puede implicar la necesidad de desarrollar programas de selección
específicos para cada ambiente particular de producción con
el fin de mantener la eficacia del mejoramiento. Sus efectos han sido poco
estudiados en características de reproducción y sobrevivencia.
En general, se considera que este tipo de efectos son importantes cuando
se comparan genotipos y ambientes extremos, como Bos indicus y Bos taurus
en ambientes templado y tropical. En ocasiones, pueden influir en el diseño
de programas de selección introduciendo errores adicionales, que
hacen necesario replicar los ambientes de prueba de los genotipos y evaluar
los animales en las condiciones reales de producción.
Nuevos avances en la tecnología reproductiva y molecular
pueden permitir perfeccionar el análisis y utilización de
las interacciones entre genes y entre los genes y el medio ambiente.
Homo economicus
A pesar de que la justificación central del mejoramiento genético animal es económica, hay muchos problemas a resolver antes de una adopción de criterios económicos completamente adecuados en el diseño de programas genéticos. En rigor, la situación ideal es una donde los objetivos para el mercado futuro se conocen con precisión. De este modo, se pueden evaluar sin error los valores económicos asociados con cada componente de la respuesta y se pueden seleccionar las características a incluir en el programa. Esto nunca es completamente cierto y es necesario recurrir a aproximaciones en presencia de incertidumbre Esto obliga a realizar análisis de sensibilidad y riesgo. Para ilustrar este tipo de problema, podemos tomar como ejemplo el énfasis que en Australia, EE.UU.. y otros países se le está dando en los programas genéticos al marmoleo de la carne de vacuno. Esto se basa en los altos precios que alcanza la carne con alto marmoleo (capas de grasa visible en el músculo) en cierta fracción del mercado de la carne en Japón. Resulta muy factible que este precio cambie con el tiempo, o que el mercado quede saturado antes de que se logre cambiar significativamente la población en este sentido. Incluso es posible que se desarrollen otros medios, diferentes de los genéticos para manipular este aspecto. Si los esfuerzos invertidos son desproporcionados con respecto del retorno esperado y el riesgo, la decisión de poner mucho énfasis en cambiar esta característica puede ser inadecuada. Esto estimula la necesidad de realizar estudios para determinar qué opciones pueden ser mas adecuadas para la industria.
Mientras que el usuario del material genético puede tener una idea más o menos clara de los objetivos de selección, es decir cuáles son las principales variables que requieren maximizar para incrementar el rendimiento económico (por ejemplo la producción de kg de cordero por oveja y por hectárea por año), para determinar cuáles son las características que se deben medir y cuáles son las ponderaciones económicas asociadas con ellas, es necesario diseñar un modelo bioeconómico del sistema de producción.
Recientemente, se han introducido aspectos sociales como criterios adicionales en la definición de objetivos de selección. En realidad estos criterios han existido siempre, pero en cierto modo no se habían realizado intentos para incorporarlos en forma sistemática en los objetivos de selección. Un ejemplo pueden ser criterios que incrementen la sustentabilidad de un sistema de producción. Se pueden desarrollar índices que combinen en una medida sintética el mérito total de los animales para un mercado en particular combinando aspectos económicos y sociales. Resulta frecuentemente preferible basar la selección en criterios económicos y utilizar aspectos complementarios como los biológicos para afinar y poner una dirección razonable al cambio genético de varias características simultáneamente.
Los objetivos biológicos pueden estar basados en la maximización de medidas físicas de la producción (por ejemplo kg de proteína por hectárea por año). Se ha argumentado que este tipo de medidas pueden ser más estables que las económicas, considerando la necesidad de incrementar la productividad a largo plazo en presencia de fluctuaciones de precios y otros componentes financieros. Una pregunta interesante es si es recomendable para cada unidad de producción seguir un programa genético que haga máximos los retornos, ponderando en forma específica las características, o bien toda la industria debe tender a utilizar un criterio uniforme de selección. Esta pregunta aún no tiene una respuesta clara. El segundo enfoque sin embargo, parece adecuado, suponiendo que los efectos del ambiente pueden ser en cierta medida optimizados y estandarizados. Esto no es siempre posible en sistemas de pastoreo o donde las fluctuaciones ambientales son parte del sistema.
Estructura de las poblaciones
En muchos casos observamos la presencia de estructuras piramidales de la población, en la que el mejoramiento fluye desde el ápice de la pirámide hacia la base. El ápice es el núcleo o grupo de animales élite. Frecuentemente este núcleo es el único grupo de la población que está en control de producción. Aquí podemos distinguir estructuras de núcleos abiertos y dispersos, como es el caso de las poblaciones de vacunos productores de leche, donde cualquier vaca registrada de alguna raza en control de producción, puede convertirse en madre de un semental de inseminación artificial y por lo tanto incorporarse a este núcleo, o núcleos cerrados como en pollos de engorde y gallinas productoras de huevo, donde los animales (genes) de los estratos inferiores nunca vuelven hacia arriba. En este tipo de estructuras puede haber estratos intermedios multiplicadores, que permiten incrementar el número de machos para ser usados como reproductores en la base. Se conoce que la base, después de un tiempo, llega a tener un mejoramiento por año similar al observado en el núcleo cuando en la base se usan machos del núcleo como reproductores. Sin embargo, existe un retraso asociado con el nivel genético, correspondiente a dos generaciones por estrato. La inseminación artificial y la transferencia de embriones fecundados desde el núcleo para ser usados en la base, son medios de reducir este retraso. Por lo anterior, la expresión económica del mejoramiento y la consanguinidad en la población base, deben ser evaluadas a nivel de la población comercial, dado que tanto la superioridad genética como la consanguinidad, se van a extender en forma diferente, dependiendo de si existen estratos multiplicadores, de si se usa IA o si el estrato comercial está basado en animales cruzados o de una sola población (ver sección siguiente). Cuando los animales de la población comercial son cruzas de poblaciones no emparentadas, la importancia práctica de la consanguinidad se reduce considerablemente. El peligro de los métodos basados en núcleos cerrados consiste en la posibilidad de que interacciones genotipo x ambiente reduzcan la respuesta genética en la población comercial cuando el núcleo es mantenido en condiciones de ambiente (alimentación, manejo, clima) diferentes a la base. Esto puede ocurrir cuando un país depende de la importación de material genético evaluado en condiciones ambientales diferentes.
Razas, cruzas y VGEs de animales cruzados
Los modelos de evaluación genética se aplican generalmente a reproductores dentro de una población, por ejemplo una línea o raza de animales. Tradicionalmente, la selección y el cruzamiento han sido consideradas como estrategias separadas para el mejoramiento genético. El uso de cruzamientos es muy común en especies productoras de carne, donde es deseable que las líneas femeninas sean fértiles y relativamente ligeras para reducir los costos de mantenimiento, mientras que los animales de engorda se espera que sean de rápido crecimiento, lo que muchas veces está asociado con una menor fertilidad y mayor peso adulto, dando mayores costos de mantenimiento en raza pura.
Una manera de combinar estas ventajas es mediante el cruzamiento de una línea (raza) ligera de hembras, con una línea (raza) de machos de rápido crecimiento, de este modo se puede optimizar el sistema, balanceando el mérito genético del crecimiento, con el de las necesidades de mantenimiento de las hembras. Cuando las hembras son cruzadas, se puede obtener mayor fertilidad y ventajas al usarlas como madres. Otro aspecto muy importante del cruzamiento, reside en el fenómeno llamado heterosis o vigor híbrido. Este fenómeno permite explotar el hecho de que diferentes alelos han sido fijados en diversos loci en razas distintas. Al cruzar dos razas, podemos esperar un incremento en la heterocigosis, es decir en la proporción de loci heterocigóticos comparados con los valores dentro de cada raza. Generalmente, este incremento en la heterocigosis se asocia a un mayor rendimiento productivo, una mayor fertilidad y una mayor tasa de supervivencia. Este vigor híbrido o heterosis se explica con teorías que postulan tanto efectos genéticos de dominancia como epistasis. A nivel teórico se supone que un animal que contiene dos tipos alternativos de genes es superior en su capacidad de adaptación al medio ambiente. La depresión observada por consanguinidad es la otra cara de la heterosis, dado que el incremento en el coeficiente de consanguinidad está asociado a una pérdida de heterocigosis.
En especies de gran fecundidad, como las aves y los cerdos, el
uso de las ventajas de los cruzamientos se basan en la cría de animales
cruzados para las granjas comerciales a partir de cruzas entre animales
de raza pura, o bien en sistemas de cruzamientos que al reemplazar las
razas de los machos utilizados en forma rotacional, permiten mantener altos
los niveles de heterosis. En bovinos y rumiantes en general, esta estrategia
no es fácil de usar por su baja fecundidad, que restringe el número
de animales cruzados comparados con el total de raza pura. Por esto, muchas
nuevas razas o poblaciones sintéticas, producto de diversos cruzamientos
entre varias razas y posterior reproducción entre sí, se
han venido desarrollado. La idea aquí es poder aprovechar la heterosis
y realizar la selección en una sola población. Asimismo,
en muchos casos se trata además de combinar características
deseables que se encuentran en poblaciones diferentes como la adaptación
a medios ambientes difíciles y la capacidad de producir con mayor
eficiencia. Un ejemplo de este tipo de animal es la popular raza Santa
Gertrudis de bovinos productores de carne, que fue desarrollada a partir
de cruzamientos entre las razas Shorthorn (Bos taurus) y Brahman
(Bos indicus). En ese caso se trató de combinar las mejor calidad
de la carne, fertilidad del Shorthorn, con la mayor tolerancia al calor
y las garrapatas del Brahman. En el caso de las poblaciones sintéticas,
una proporción de la heterosis es retenida comparada con el nivel
exhibido por las cruzas entre dos razas puras. Dos principales problemas
que han limitado la difusión de razas sintéticas, son su
menor aceptación por parte de los granjeros y por el otro, en muchos
casos lo pequeño de las poblaciones, que ha llevado a incrementos
notables en la consanguinidad, con lo que se pueden perder las ventajas
de la heterosis y ofrece pocas oportunidades para lograr mejoras importantes
por selección.
 |
| Figura 1. La raza sintética Australian Friesian Sahiwal. Esta raza está siendo desarrollada en Australia por el Gobierno de Queensland para ser usada en áreas tropicales. La raza fue creada a partir de Sahiwal, una variedad lechera de Cebú de Pakistán y la Holstein-Friesian Australiana. Desde la decada de 1960, cuando comenzó la investigación sobre esta raza, se han logrado notables progresos hacia el objetivo de combinar reistencia a las garrapatas y tolerancia al calor con consistentes niveles de producción de leche y fertilidad. Actualmente ha sido extensamente probada en áreas tropicales y sub-tropicales de Australia. Bajo esas condiciones, produce aproximadamente 15% mas leche que la Holstein-Friesian. Nota: Esta foto e información se proporciona por cortesía de Meat and Livestock Australia Locked Bag 991, North Sydney 2059, Australia. |
Una idea más nueva y posiblemente más práctica para explotar al mismo tiempo las diferencias entre y dentro de razas, consiste en el cálculo de VGEs de animales cruzados. Esto se puede lograr corrigiendo para diferencias entre las medias, de cada raza y para la heterosis. De este modo se pueden obtener comparaciones entre y dentro de razas, que permiten seleccionar animales dentro y a través de poblaciones. Dado que algunos efectos son estimables a partir de las bases de datos existentes, es posible aprovechar estas ideas para hacer evaluaciones simultáneas de los valores genéticos dentro de raza, los efectos de raza y de heterosis, con la idea de optimizar el sistema. En muchos casos sin embargo, es necesario diseñar cruzas específicas para obtener la información requerida.
Optimicemos
Durante un tiempo, el affaire entre los modelos lineales, que daban predicciones insesgadas de los valores genéticos y la estimación de los parámetros requeridos para su uso en poblaciones de una raza, consumió una gran parte de los esfuerzos de los genetistas de animales. La solución de algunos importantes problemas quedó pendiente hasta hace muy poco tiempo. En efecto, la optimización de los programas genéticos que involucra aspectos tales como la maximización de la respuesta mientras se mantienen restringidos los costos del programa asociados al uso de cada reproductor, al uso de tecnologías reproductivas y el incremento en la consanguinidad, han recibido menos atención.
No es obvio por ejemplo cuáles y cuántos animales utilizar en un programa genético, ni cuanta progenie obtener de cada uno, o si es económicamente viable obtener embriones de una vaca en particular con OMTE. Contestar este tipo de preguntas con precisión no ha sido posible sino hasta ahora, con el desarrollo de metodologías de maximización muy flexibles basadas en algoritmos genéticos. Recientemente, se han desarrollado y comenzado a aplicar en la industria estrategias llamadas tácticas, que permiten optimizar problemas complejos usando como entradas índices económicos obtenidos con los VGEs, el pedigrí y los costos de reproducción, en cada momento de decisión; por ejemplo cada año antes de realizar los apareamientos. En este tipo de estrategias, que maximizan una función que involucra la respuesta genética y otros componentes, pueden ser instrumentados mediante la selección y asignación de apareamientos.
Algunos trabajos pioneros están siendo desarrollados en este campo, considerando estructuras de apareamiento óptimas para el conjunto de machos y hembras de la población. Las soluciones son obtenidas con el uso de algoritmos genéticos, que permiten la maximización de la respuesta para niveles predefinidos de consanguinidad. Cuando se trata de poblaciones cruzadas, se pueden diseñar estrategias específicas de maximización de los efectos de heterosis.
Uso de tecnología reproductiva y molecular
Mientras las técnicas moleculares ofrecen una serie importante de posibilidades para la mejora genética animal, la materialización de estas expectativas requiere de la solución de un número importante de problemas técnicos para aprovechar toda la información disponible de un modo eficiente, reducir los costos de generar información genómica y obtener estimaciones confiables de los efectos de genes mayores con efectos en características productivas (QTLs) y de la aplicación de la selección asistida por marcadores (SAM) y la información genómica en general para la mejora animal.
Un uso racional de las metodologías moleculares requiere el uso óptimo de los métodos tradicionales de selección. El máximo provecho puede ser obtenido cuando estas técnicas son utilizadas en forma integrada con tecnologías reproductivas como la inseminación artificial, la colección y producción in vitro de embriones para acelerar el cambio genético.
Las características en las cuales la aplicación
de la SAM puede ser más efectiva son aquellas que se miden tardíamente
en la vida del animal, o que son controladas por pocos pares de alelos.
El primer ejemplo corresponde a la longevidad y las características
de composición corporal en animales productores de carne, el segundo,
a la resistencia a ciertas enfermedades o defectos de herencia simple.
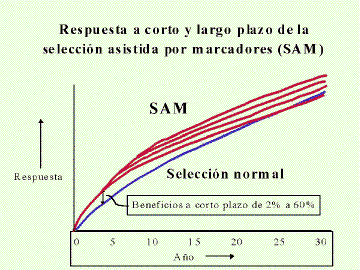 |
| Figura 2. Respuesta a corto y largo plazo de la selección asistida por marcadores de ADN (SAM), las líneas rojas muestran el avance posible usando SAM comparada a selección convencional en una característica cuantitativa. El efecto de la SAM será incrementar la respuesta a corto plazo (Reproducido con autrorización de: Kinghorn, B.P. (1998). Managing genetic change under operational and cost constraints. 36th National Congress of the South African Association of Animal Science. University of Stellenbosch 5-8 April. pp 9-16). |
La selección puede incorporar el llamado análisis
de segregación para detectar y usar posibles genes de efectos
mayores en la selección. La principal ventaja de los métodos
moleculares sobre el análisis de segregación, es la posibilidad
de evaluar el efecto simultáneo de varios QTLs
sobre las características de importancia económica, y en
el futuro incrementar su precisión y la complejidad de los modelos
de acción génica involucrados, por ejemplo QTLs con alelos
múltiples. Las tendencia actual indica que los dos métodos
se integrarán en el futuro en los programas de mejora genética
animal con métodos basados en modelos que contienen el efecto de
los genes mayores y herencia poligénica (modelos de herencia mixta).
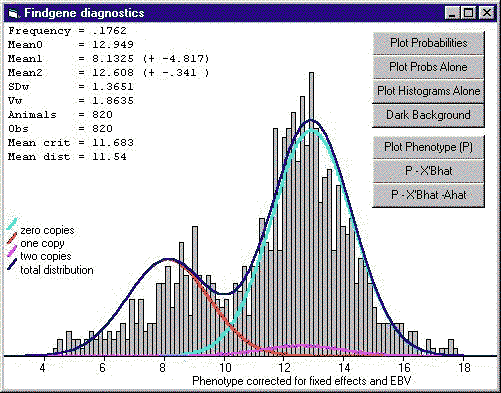 |
| Figura 3. Ejemplo de utilización del sotware Findgene para realizar análisis de segregación junto a un modelo lineal para detectar un gene de efecto mayor en el fenotipo de una característica cuantitativa. Se observa la distribución total de fenotipos (linea azul) y las distribuciones de cada genotipo para el gene mayor, representadas por las líneas roja, azul claro y púrpura. Las barras grises muestan la distribución observada de los fenotipos corregidos para efectos ambientales y de los poligenes (Kinghorn, B.P., 1997). |
Es preciso puntualizar que probablemente pasarán muchos años antes de que las tecnologías reproductivas y moleculares revolucionen el mejoramiento animal al grado de hacer innecesario el control de producción, si bien esto resulta conceptualmente posible con la reproducción de células en cultivo usando selección puramente molecular de las variantes deseables, seguidas de clonación y multiplicación de la variante genética deseada. El conocimiento que permita este desarrollo (si este es posible) se tendrá que generar combinando información molecular con información fenotípica. Por el otro lado, es poco probable que el control de producción se abandone completamente, dado que se tiene que seguir manteniendo como un medio de control de que las expectativas y predicciones se están cumpliendo en la práctica.
Las técnicas moleculares y otras técnicas analíticas pueden posibilitar la medición de nuevas características en los programas genéticos que estén relacionadas en forma más estrecha con los objetivos del programa de mejoramiento.
Conclusiones
La evaluación de animales en base de datos de mediciones (fenotipo) y pedigrí para calcular los valores genéticos estimados (VGEs), seguirán en el futuro siendo la base del mejoramiento animal. Se esperan mejoras en los métodos para ponderar económicamente diferentes características para la selección, para maximizar la respuesta a los programas genéticos, controlando costos y consanguinidad con nuevas herramientas de análisis y optimización de problemas complejos y no-lineales como los algoritmos genéticos y la inteligencia artificial. Se espera la incorporación de nuevas características que se relacionen en forma mas directa con los objetivos de selección.
Otros avances son la generalización de métodos para evaluar genéticamente poblaciones compuestas por diversas razas y cruzas y el desarrollo de métodos para incorporar la información genómica (marcadores moleculares de ADN) y usar nuevas técnicas reproductivas basadas en la manipulación de la fisiología reproductiva de los animales, gametos y embriones en el diseño de programas más eficientes de mejora animal.
Posiblemente los desarrollos de la biología molecular puedan ayudar a un mejor entendimiento de fenómenos aún no bien comprendidos como la epistasis (interacciones entre alelos en distintos loci) y las interacciones genotipo x ambiente, al posibilitar el análisis del efecto de genes específicos en caracteres de herencia compleja como los involucrados en el mejoramiento animal.
Referencias
Bibliografía
Ashwell, M. S., RexroadJr., C. E., Miller, R. H., Van Raden, P.M. and Da, Y. (1997). Detection of loci affecting milk production and health traits in an elite US Holstein population using microsatellite markers. Animal Genetics 28:216-222.
Cameron, N. D. (1997). Selection indices and prediction of genetic merit in animal breeeding. CAB International. Wallingford, UK.
Falconer, D. S. (1989). Introduction to quantitative genetics. 3rd ed. Longman, Essex, UK.
Goddard, M. E. (1992). A mixed model for analyses of data on multiple genetic markers. Theor. Appl. Genet. 83: 878-886.
Goddard, M. E., Barwick, S. A. and Kinghorn, B. P. (1998). Breeding objectives for meat animals: development of a profit function. Animal Production in Australia 22: 90-94.
Groen AF, Steine T., Colleau J. J., Pedersen J., Prybyl J., Reinsch N. (1997). Economic values in dairy cattle breeding, with special reference to functional traits. Report of an EAAP-working group. Livestock Production Science 49:1-21.
Haley, C. S. (1995). Livestock QTLs ? bringing home the bacon? Trends Genet. 11(12):488-492.
Henderson, C. R. (1984). Applications of linear models in animal breeding. University of Guelph, Ontario, Canada
Hill WG. (1971). Investment apprisal for national breeding programmes. Animal Production 3:37-50.
Janss, L. L. G., Thompson, R. and van Arendonk, J. A. M. (1995). Application of Gibbs sampling for inference in a mixed major gene-polygenic inheritance model in animal populations. Theor. Appl. Genet. 91: 1137-1147.
Keown J. F., Montaldo H., Van Vleck L. D., Van Tassell, C. P. (1998). Economic responses and risk from use of selected Holstein sires in Italy, Mexico, The Netherlands and USA. Proceedings of the 6th World Congress on Genetics Applied to Livestock Production; 1998 January 11-16; Armidale, NSW, Australia 23:327-330.
Kinghorn, B. P. (1982) Genetic effects in crossbreeding. I. Models of Merit. Z. Tierzüch. Züchtgs-biol. 99: 59-68.
Kinghorn, B. P. (1997). A view on future developments in animal breeding. Slides from a 1997 A.B. Chapman Lecture, University of Wisconsin, Madison. http://ops.agsci.colostate.edu/~bkinghor/future/ppframe.htm
Kinghorn, B. P., Grundy, B., Shepherd, R. K., and Woolliams, J. A. (1997) Use of a stochastic algorithm to make selection and reproduction decisions under operational and cost constraints http://metz.une.edu.au/~bkinghor/SelRep/2Stage_Matesel2.html
Kinghorn, B. P., van Arendonk, J. A. M. and Hetzel, J. (1994). Detection and use of major genes in animal breeding. AgBiotech News and Information 6(12): 297N-302N
Lynch, M. and Walsh, B. (1998). Genetic and analysis of quantitative traits. Sinauer, Sunderland, USA
Meuwissen, T. H. E. and van Arendonk, J. A. M. (1992). Potential improvements in rate of genetic gain from marker assisted selection in dairy cattle breeding schemes. J. Dairy Sci. 75: 1651-1659.
Meuwissen, T. H. E. and Goddard, M. E. (1996). The use of marker-haplotypes in animal breeding schemes. Genet. Sel. Evol. 28:161-176.
Miller, S. P. and Goddard, M. E. (1998). The super breed approach to modelling additive and non-additive genetic effects between and within breeds in multi-breed evaluations. Proceedings of the 6th World Congress on Genetics applied to livestock production, Armidale, NSW 23:97-100 (1998.
Montaldo, H. (1993). Biotecnología y mejoramiento genético animal en México. En: (M. Arenas, L. F. Bojalil y L. Hernández Comp. Las profesiones en México: Agronomía, Medicina Veterinaria y Zootecnia, Universidad Autónoma Metropolitana-Xochimilco, Universidad de Colima, México, D. F. pp 90-108)
Montaldo, H. and Meza-Herrera, C. A. 1998. Use of molecular markers and major genes in the genetic improvement of livestock. Electronic Journal of Biotechnology 1(2) http://ejb.ucv.cl/content/vol1/issue2/full/4/
Montaldo, H., Keown, J. F. and Van Vleck, L. D. (1998). Effect of in vitro embryo production and sexed semen in dairy MOET nucleus systems. Proceedings of the 6th World Congress on Genetics applied to livestock production, Armidale, NSW 25:443 (1998).
Montgomery, G. W. and Kinghorn, B. P. (1997). Recent developments in gene mapping and progress toward marker-assisted selection in sheep. Australian Journal of Agricultural Research 48: 729-741.
Mrode, R. A., 1996. Linear Models for the prediction of animal breeding values. CAB International, Wallingford, UK. pp. 187.
Nicholas, F. W. (1996). Introduction to veterinary genetics. Oxford University Press, Oxford, UK.
Ruane, J., G. Klemesdal and E. Sehested. (1997). Views on the potential impact of cloning on animal breeding and production. Acta Agric. S-cand. 47: 209-212.
Weller J. I. (1994). Economic aspects of animal breeding. 1st ed. London, Chapman and Hall.
Wakayama, T., A. C. F. Perry, M. Zuccotti, K. R. Johnson and R. Yanagimachi. (1998). Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 394: 369-373.
Woodward, B. W., Du, F.-X., Montaldo, H., Andersen, K. J. and DeNise, S. K. (1998). Preliminary evidence for major genes controlling beef carcass traits in Limousin cattle. Proc. of the 6th Word Congress on Genetics Applied to Livestock Production, Armidale, NSW. Vol. 25: 157-160.
